StW 573, a hominin skeleton more palatably nicknamed “Little Foot,” made its big debut last week:

Ron Clarke showcases the lovingly-excavated skeleton (Photo credit: AP/Themba Hadebe)
The skeleton is remarkable in that it is the most complete australopithecine individual ever discovered, and is among the most complete in the entire hominin fossil record. Below I’ve compared it to the most complete Australopithecus afarensis (KSD-VP-1/1 and AL 288-1), A. africanus (StW 431 and Sts 14), and A. sediba (MH1-2); the Dikika infant would be a neat comparison, too, but I don’t know of any photos of its bones nicely laid out. The other skeletons are practically naked (or dismembered) compared to Little Foot.

Little Foot (red) compared with other australopithecine skeletons. Images not to scale! (Photo credit: The Internet!)
Beyond it’s completeness, the other parts of story of Little Foot are equally fascinating – from its discovery based on already-known fragments to the possibility that it is older than “Lucy” (AL-288). Ron Clarke has painstakingly and I’d say very successfully removed the skeleton from the hard breccia in which the fossils were encased. Having spent the better part of the past two decades with the skeleton, he has argued that Little Foot represents a second hominin species at Sterkfontein, Australopithecus prometheus (Clarke 2013), the species to which hominin fossils at Makapansgat were originally attributed (Dart 1948). With the unveiling of the skeleton, I’d guess that in the coming years we’ll see renewed investigations into the number of species at Sterkfontein, and the general comparison between hominins from there and Makapansgat.
From pictures in the media releases, we can see a few things that weren’t known from previous publications. I’ll outline a few here, but emphasize that these are only superficial observations and will need to be borne out by further research.

“EXPELLIARMUS”
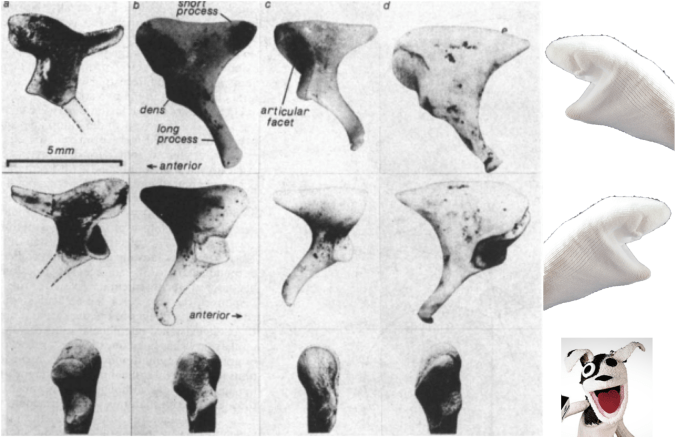
At the top of the trunk, the cervical vertebra seems to have a fairly wide spinal canal, a human-like ‘bulging’ which Meyer and Hausler (2015) suggest might reflect innervation of highly manipulative hominin hands.

Close up of the skull and upper trunk of StW 573, highlighting the cervical vertebral canal (white arrow) and first rib (orange arrow). Original photo credit: AP//Themba Hadebe.
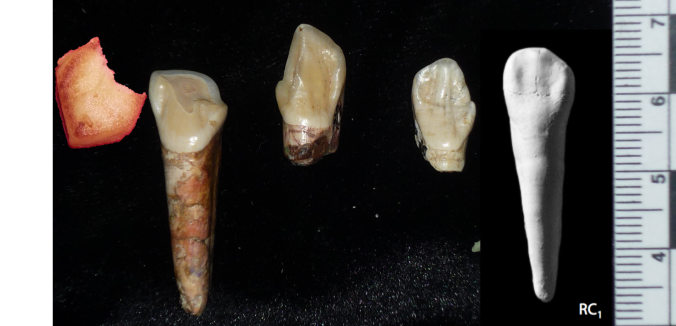
In addition, the first rib may be relatively long front-to-back (as opposed to wide side-to-side), possibly indicating a more barrel-like chest than in other early hominins; the angle of the photo and the clear break between the proximal and distal portions, however, makes this unclear.

Hominin first ribs/bacon, with StW 573 on the far right. Not to scale! Modified from this post.
The distal forelimb (i.e., radius and ulna) are not as elongated as in apes, but the femur is not as elongated as in the genus Homo. From the pictures, the femur neck appears short like in humans, not as elongated as is characteristic for australopiths and early Homo.

Limb proportion comparison. Humerus (top row), radius & ulna (middle), and femur (bottom). Image modified from Asfaw et al. (1999). StW 573 scaled to same humerus length as the human. Note also that all bones are from the right except the StW 573 upper limb.
The apparently short femur neck, similar to humans, contrasts with the wide, flaring ilium of the pelvis. This appears fairly flat, short and wide (Australopithecus af) compared to modern humans’ more strongly curved ilium. But this inference is just from a picture and it’s likely that the fossil needs a bit of reconstruction to uncover the true anatomy.

StW 573 pelvis (left) compared with Sts 14 (A. africanus, middle) and SH pelvis 1 (archaic Homo, right). Sts 14 modified from Berge & Goularis (2010) and SH pelvis 1 from Bonmati et al. (2010).
I’d like to emphasize that these are just first impressions based on press release photos, and actual analysis of the skeleton are necessary to tell if these impressions are correct. As could be expected, the skeleton as a whole looks typically australopithecine, although the short femur neck may be a bit different. As 2017 draws to a close, let’s hope 2018 sees the testing of these predictions.
References
Asfaw B. et al. 1999. Australopithecus garhi: a new species of early hominid from Ethiopia. Science 284: 629-635.
Berge C and Goularis D. 2010. A new reconstruction of Sts 14 pelvis (Australopithecus africanus) from computed tomography and three-dimensional modeling techniques. Journal of Human Evolution 58: 262-272.
Bonmati A. et al. 2010.Middle Pleistocene lower back and pelvis from an aged human individual from the Sima de los Huesos site, Spain. Proceedings of the National Academy of Sciences 107: 18386-18391.
Clarke RJ. 2013. Australopithecus from Sterkfontein Caves, South Africa. In The Paleobiology of Australopithecus, Reed et al., eds. Dordrecht: Springer Science+Business.
Dart R. 1948. The Makapansgat proto-human Australopithecus prometheus. American Journal of Physical Anthropology 6: 259-284.
Meyer M. and Hausler M. 2015. Spinal cord evolution in early Homo. Journal of Human Evolution 88: 43-53.