The Fall semester here at Vassar kicks off next week, and so of course a new fossil discovery is published this week that threatens to upend my course plans and throw my syllabi into disarray. Haile-Selassie and colleagues report a very well-preserved hominin cranium, from the Woranso-Mille region of Ethiopia and dating to 3.8 million years ago. The new cranium shares features with Australopithecus anamensis, a species previously mainly known through jaws and teeth. The fossil is therefore really important since it puts a face to the species’ name, and it is the oldest relatively complete Australopithecus cranium known. When I showed a picture of the fossil to my wife, who is not a paleoanthropologist, all she said was that it looked like the face of a dog who got stung by a bee.
The new A. anamensis fossil MRD-VP-1 (left), and a dog that lost a fight with a bee. Fossil photo from the Smithsonian‘s coverage.
The big buzz in many news stories about the fossil (for example, Nature, ScienceNews, etc.) is that it rewrites an evolutionary relationship early in human history, with Australopithecus anamensis no longer the ancestor of A. afarensis, but rather the two being contemporaries. That idea is based on a 3.9 million year old frontal bone attributed to A. afarensis from a site called Belohdelie, also in Ethiopia (Asfaw, 1987): basically, the new A. anamensis cranium reveals a hominin with a narrow frontal region of the brain, which lived 100,000 later than A. afarensis with a relatively expanded frontal region:
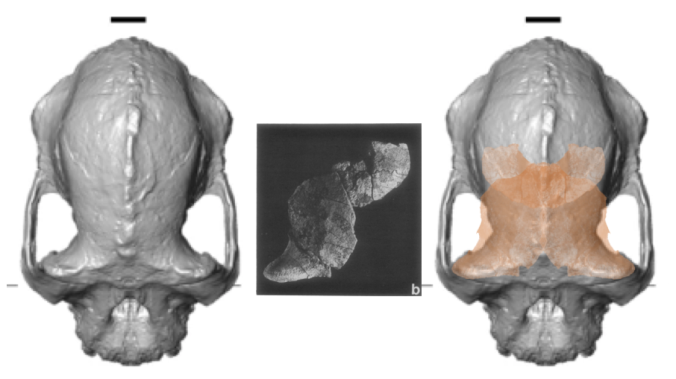
Top views of the reconstructed A. anamensis cranium (left), and the Belohdelie frontal (center), and my crappy photoshopped overlay of Belohdelie on A. anamensis (right). Images not to scale.
The lede, “human evolutionary tree messier than thought,” is not terribly interesting or compelling since it seems to characterize most fossil discoveries over the past several years. And in this case I don’t know how well supported the argument is, since the trait in question (narrow frontal region of the braincase or “post-orbital constriction”) can vary dramatically within a single species. The image below is from the paper itself—compare the difference in “postorbital constriction index” (left graph) between the new A. anamensis cranium (MRD) and A. afarensis (in blue). Both sets of fossils fall within the range of chimpanzees (P. troglodytes), and note the great range of variation within gorillas (G. gorilla).
Part of Figure 3 from the paper by Haile-Selassie and colleagues. On the top is a view from above of fossil humans: Sahelanthropus tchadensis, Ardipithecus ramidus, the new A. anamensis, A. afarensis, and A. africanus. Below the graphs show how species differ in narrowing of the frontal (left) and length of the skull (right).
What I find most interesting about the new find is the great front-to-back length of the cranium—check out how long and narrow the brain-case is of the fossil compared with the later hominins to the right. This is an interesting similarity with the much earlier (6 million years ago) Sahelanthropus tchadensis, which is the left-most fossil in the figure. It makes me really curious to see the brain endocast of A. anamensis and the Sahelanthropus cranium—what was brain shape like for these ancient animals, and what does that mean for the earliest stages of human brain evolution? The Sahelanthropus endocast was meticulously reconstructed and analyzed by Thibaut Bienvenu (dissertation here), who found it to be somewhat like gorillas in overall shape (but with a more hominin-like placement of the foramen magnum). Of course, the outside of the skull doesn’t necessarily tell us what the underlying brain shape was like. Haile-Selassie and colleagues report that they made a virtual reconstruction of the A. anamensis endocast, so hopefully we’ll get to pick its brain soon.
As we’re wrapping up what may be the worst year in recent global memory, especially geopolitically, let’s take a moment to review some more positive things that came up at Lawnchair in 2016.
Headed home
Alternate subtitle: Go West
This was a quiet year on the blog, with only 18 posts compared with the roughly thirty per year in 2014-2015. The major reason for the silence was that I moved from Kazakhstan back to the US to join the Anthropology Department at Vassar College in New York. With all the movement there was less time to blog. Much of the second half of 2016 was spent setting up the Biological Anthropology Lab at Vassar, which will focus on “virtual” anthropology, including 3D surface scanning…
Cast of early Homo cranium KNM-ER 1470 and 3D surface scan made in the lab using an Artec Spider.
… and 3D printing.
A gibbon endocast, created from a CT scan using Avizo software and printed on a Zortrax M200.
This first semester stateside I reworked my ‘Intro to Bio Anthro’ and ‘Race’ courses, which I think went pretty well being presented to an American audience for the first time. The latter class examines human biological variation, situating empirical observations in modern and historical social contexts. This is an especially important class today as 2016 saw a rise in nationalist and racist movements across the globe. Just yesterday Sarah Zhang published an essay in The Atlantic titled, “Will the Alt-right peddle a new kind of racist genetics?” It’s a great read, and I’m pleased to say that in the Race class this semester, we addressed all of the various social and scientific issues that came up in that piece. Admittedly though, I’m dismayed that this scary question has to be raised at this point in time, but it’s important for scholars to address and publicize given our society’s tragically short and selective memory.
So the first semester went well, and next semester I’ll be teaching a seminar focused on Homo naledi and a mid-level course on the prehistory of Central Asia. The Homo naledi class will be lots of fun, as we’ll used 3D printouts of H. naledi and other hominin species to address questions in human evolution. The Central Asia class will be good prep for when I return to Kazakhstan next summer to continue the hunt for human fossils in the country.
Osteology is still everywhere
A recurring segment over the years has been “Osteology Everywhere,” in which I recount how something I’ve seen out and about reminds me of a certain bone or fossil. Five of the blog 18 posts this year were OAs, and four of these were fossiliferous: I saw …
And a Homo erectus cranium on a Bangkok sidewalk. As I’m teaching a fossil-focused seminar next semester, OA will probably become increasingly about fossils, and I’ll probably get my students involved in the fun as well.
New discoveries and enduring questions
The most-read post on the blog this year was about the recovery of the oldest human Nuclear DNA, from the 450,000 year old Sima de los Huesos fossils. My 2013 prediction that nuclear DNA would conflict with mtDNA by showing these hominins to be closer to Neandertals than Denisovans was shown to be correct.
These results are significant in part because they demonstrate one way that new insights can be gained from fossils that have been known for years. But more intriguingly, the ability of researchers to extract DNA from exceedingly old fossils suggests that this is only the tip of the iceberg.
The comparison between monkey-made and anthropogenic stone tools drives home the now dated fact that humans aren’t the only rock-modifiers. But the significance for the evolution of human tool use is less clear cut – what are the parallels (if any) in the motivation and modification of rocks between hominins and capuchins, who haven’t shared a common ancestor for tens of millions of years? I’m sure we’ll hear more on that in the coming years.
In the case of whether Neandertal brain development is like that of humans, I pointed out that new study’s results differ from previous research probably because of differences samples and methods. The only way to reconcile this issue is for the two teams of researchers, one based in Zurich and the other in Leipzig, to come together or for a third party to try their hand at the analysis. Maybe we’ll see this in 2017, maybe not.
There were other cool things in 2016 that I just didn’t get around to writing about, such as the publication of new Laetoli footprints with accompanying free 3D scans, new papers on Homo naledi that are in press in the Journal of Human Evolution, and new analysis of old Lucy (Australopithecus afarensis) fossils suggesting that she spent a lifetime climbing trees but may have sucked at it. But here’s hoping that 2017 tops 2016, on the blog, in the fossil record, and basically on Earth in general.
These headlines, each saying something slightly different, are referring to a study by Indjeian and colleagues published in Cell. Researchers identified a stretch of DNA that is highly conserved across mammals, or in other words, it is very similar between very different organisms. In humans, however, this conserved region is actually missing (“hCONDEL.306”):
Fig. 4A from Indjeian et al. 2016. A stretch of DNA on Chromosome 8, “hCONDEL.306,” is very similar between chimpanzees, macaque monkeys, and mice, but is completely missing in humans (as is another stretch, hCONDEL.305).
That a stretch of DNA should be highly conserved across diverse animal groups suggests purifying natural selection has prevented any mutations from occurring here – alterations to this stretch of DNA negatively affected fitness. But that humans should be missing such a highly conserved region suggests that this deletion came under positive natural selection at some point in human evolution. This strategy, of seeking stretches of DNA that are similar between many animals but very different in humans, has led to the identification of hundreds of genetic underpinnings of human uniqueness. Some of these, such as the case in question, involve deleted sequences and have been termed “hCONDELs,” for “regions with high sequence conservation that are surprisingly deleted in humans” (McLean et al., 2011: 216). Others involve the accumulation of mutations where other animals show few or none (e.g., HACNS1; Prabhakar et al. 2008). In many (most?) cases these are “non-coding” sequences of DNA.
How can “non-coding” DNA help make humans upright?
As was predicted 30 years ago (King and Wilson, 1975), what makes humans different from other animals isn’t so much in the protein-coding DNA (the classical understanding of the term, “genes”), but rather in the control of these protein-coding genes. “Non-coding” means that a stretch of DNA may get transcribed into RNA but is not then translated into proteins. But even though these sequences themselves don’t become anything tangible, many are nevertheless critical in regulating gene expression – when, where and how much a gene gets used. It’s wild stuff. Indeed, “Many human accelerated regions are developmental [gene] enhancers” (Capra et al., 2013).
In the present case, hCONDEL.306 refers to the human-specific deletion of a developmental enhancer located near the GDF6 gene, which is a bone morphogenetic protein. The major finding of the paper, as stated succinctly in the Highlights title page, is that “Humans have lost a conserved regulatory element [hCONDEL.306] controlling GDF6 expression…. Mouse phenotypes suggest that [this] deletion is related to digit shortening in human feet.”
How do they link this “gene tweak” to digit shortening?
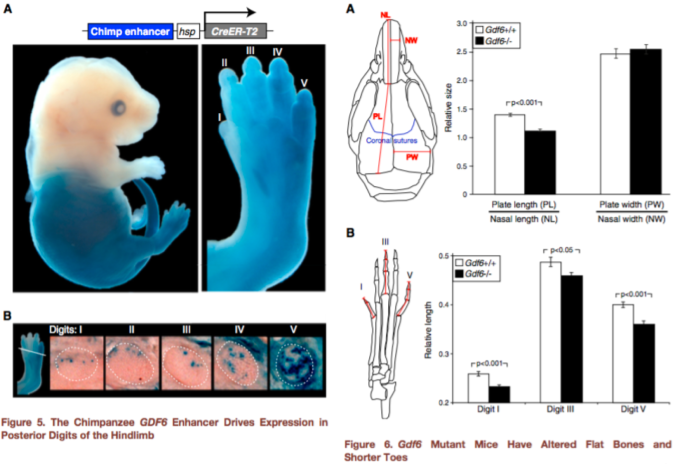
Since humans have lost this gene enhancer that is highly conserved in other mammals, Indjeian and team reasoned that the chimpanzee DNA sequence associated with this deletion, retaining the enhancer sequence, is likely the ancestral condition from which the human version evolved. They inserted the chimpanzee version into mouse embryos and watched what happened as they developed. The enhancer was only active in the mice’s back legs, specifically in the cartilage that would later become the lateral toe bones and cells that would become a muscle of the big toe (abductor hallucis). These are areas where humans and chimpanzees differ: our lateral toes are shorter than chimps’, and we only have one abductor hallucis muscle whereas chimpanzees have an additional, longer abductor hallucis (Aiello and Dean, 2002). So, we’re on our way to seeing how hCONDEL.306 might relate to our big toe or upright walking, as the headlines say.
But this still doesn’t explain how this deletion affects GDF6 gene expression, and therefore what this does for our feet. Pressing onward, the scientists compared the size of certain bones in mice with a normal Gdf6 gene, and those in which the Gdf6 gene was completely turned off (or “knocked out”). The Gdf6 knock-out mice had shorter lateral toe bones than regular mice, but they also had shorter big toes as well – the previous experiment staining mouse embryos showed the ancestral enhancer was expressed more in the latter toes, not so much the big toe.
Figures 5-6 from Indjeian et al. (2016) sum up the findings. Figure 5 (left) shows that the ancestral version of the GDF6 enhancer (blue staining) is most strongly expressed in the lower half of the body, especially the fifth toe bone. Figure 6 (right) shows that a lack of Gdf6 expression (black bars) results in shorter skull and toe bones. Combining these findings, humans lack a gene enhancer associated with the development of long lateral toes.
hCONDEL.306 doesn’t completely turn off GDF6, so this second experiment doesn’t really tell us exactly what the hCONDEL does. But the results are highly suggestive. Indjeian and team showed that Gdf6 affects toe length, among other skeletal traits, in mice. The ancestral enhancer that humans are missing seems to affect GDF6 activity in the leg/foot only. This illustrates a mechanism of modularity – as the authors state, “Loss of this enhancer would thus preserve normal GDF6 functions in the skull and forelimbs, while confining any … changes to the posterior digits of the hindlimb.” In other words, developmental enhancers allow different parts of the body to evolve independently despite being made by some of the same genes (such as GDF6).
As with any good study, results are intriguing but they raise more questions for future studies. The researchers conducted two experiments to investigate the function of hCONDEL.306: first putting the chimp version in mouse embryos to see where the ancestral enhancer is expressed, and then turning off Gdf6 completely in mice to see what happens. A more direct way to see what hCONDEL.306 does might be to put a longer stretch of the human sequence surrounding GDF6 containing (or rather missing) the ancestral enhancer into mouse embryos. I’m not a molecular biologist so maybe this isn’t possible. But this is important because the ancestral (chimpanzee) enhancer appeared to be most strongly expressed in the little toe, but of course this isn’t our only toe that is short compared to chimps. Similarly, how hCONDEL.306 relates to the abductor hallucis muscle remains in question – does it reduce the size of the intrinsic muscle present in both humans and chimps, or does it prevent development of the longer muscle that chimps have but we lack? We can expect to find hCONDEL.306 in the genomes of Neandertals (and Denisovans?), since they also have short toes, but what would it mean if they retained the ancestral enhancer?
So how does this gene tweak help with upright walking?
This is a really cool paper with important implications for human evolution, but something seems to have been lost in translation between the paper and the headlines (the news pieces themselves are good, though). The upshot of the study is that humans lack a stretch of non-coding DNA, which in chimpanzees (or chimp-ified mice) promotes embryonic development of the lateral toes and a big toe muscle. This may be a genetic basis for at least some aspects of our unique feet that have evolved under natural selection for walking on two legs.
But the headlines misrepresent this result, with words like “led to,” “allowed,” and “caused,” especially when these are followed by “big toe” or “upright walking.” hCONDEL.306 doesn’t really have anything to the big toe bone itself, although it might relate to a muscle affecting our big toe. The only sense in which the “Gene tweak led to humans’ big toe” (first title above) is that hCONDEL.306 might be responsible for our short lateral toes, which make our first toe look big by comparison. The other headlines are misleading since we know from fossil evidence that hominins walked upright long before we have evidence for short toes:
These little piggies get none. Fourth toe bones of living apes and humans (left) and (probable) hominins from 3-5 million years ago (right). I did my best to get all images to scale.
“Epigenetic,” from the fourth article headline, is simply wrong. Modern day epigenetics is a field concerned with the chemical alterations to the structure of DNA. Even the broad concept of epigenetic as originally devised by Conrad Waddington was about how environments (cellular or outside the body) influence development.
It’s hard to fit all the important and interesting information from scientific papers into news headlines. Still, it would be good if headlines more accurately portrayed scientific findings, especially avoiding such definitive verbs as “caused.” Especially in the realm of biology, people should know that there’s a lot that we still don’t know, that there’s lots more important work left to be done.
References
Aiello and Dean, 2002. Human Evolutionary Anatomy. Academic Press.
Capra et al., 2013. Many human accelerated regions are developmental enhancers. Philosophical Transactions of the Royal Society B 368: 20130025.
Indjeian et al. 2016. Evolving new skeletal traits by cis-regulatory changes in bone morphogenetic proteins. Cell http://dx.doi.org/10.1016/j.cell.2015.12.007
King and Wilson, 1975. Evolution at two levels in humans and chimpanzees. Science 188: 107-116 DOI: 10.1126/science.1090005
McLean et al., 2011. Human-specific loss of regulatory DNA and the evolution of human-specific traits. Nature 471: 216-219.
Prabhakar et al., 2008. Human-specific gain of function in a developmental enhancer. Science 321: 1346-1350.
Holy crap 2015 was a big year for fossils. And how fortuitous that 2016 begins on a Fossil Friday – let’s recap some of last year’s major discoveries.
Homo naledi
Some Homo naledi mandibles in order from least to most worn teeth.
The Homo naledi sample is a paleoanthropologist’s dream – a new member of the genus Homo with a unique combination of traits, countless remains belonging to at leasta dozen individuals from infant to old adult, representation of pretty much the entire skeleton, and a remarkable geological context indicative of intentional disposal of the dead (but certainly not homicide, grumble grumble grumble…). The end of 2015 saw the announcement and uproar (often quite sexist) over this amazing sample. You can expect to see more, positive things about this amazing animal in 2016.
We’ll be presenting a bunch about Homo naledi at this year’s AAPA meeting in Hotlanta. I for one will be discussing dental development at Dinaledi- here’s a teaser:
As long as we’re talking about the AAPA meetings, my colleague David Pappano and I are organizing a workshop, “Using the R Programming Language for Biological Anthropology.” Details to come!
Lemur graveyard
Homo naledi wasn’t the only miraculously copious primate sample announced in 2015. Early last year scientists also reported the discovery of an “Enormous underwater fossil graveyard,” containing fairly complete remains of probably hundreds of extinct lemurs and other animals. As with Homo naledi, such a large sample will reveal lots of critical information about the biology of these extinct species.
Australopithecus deyiremeda
Extended Figure 1h from Haile-Selassie et al. (2015), compared with Demirjian developmental stages 6-8 . While the M1 roots look like stage 8 (complete), M2 looks like stage 7 (incomplete).
We also got a new species of australopithecus last year. Australopithecus deyiremeda had fat mandibles, a relatively short face (possibly…), and smaller teeth than in contemporaneous A. afarensis. One tantalizing thing about this discovery is that we may finally be able to put a face to the mysterious foot from Burtele, since these fossils come from nearby sites of about the same geological age. Also intriguing is the possible evidence, based on published CT images (above), that A. deyiremeda had relatively advanced canine and delayed molar development, a pattern generally attributed to Homo and not other australopithecines (if this turns out to be the case, you heard it here first!).
Lomekwian stone tool industry
3D scan and geographical location of Lomekwian tools. From africanfossils.org.
Roughly contemporaneous with A. deyiremeda, Harmand et al. (2015) report the earliest known stone tools from the 3.3 million year old site of Lomekwi 3 in Kenya. These tools are a bit cruder and much older than the erstwhile oldest tools, the Oldowan from 2.6 million years ago. These Lomekwian tools, and possible evidence for animal butchery at the 3.4 million year old Dikika site in Ethiopia (McPherron et al. 2010; Thompson et al. 2015), point to an earlier origin of lithic technology. Fossils attributed to Kenyanthropus platyops are also found at other sites at Lomekwi. With hints at hominin diversity but no direct associations between fossils and tools at this time, a lingering question is who exactly was making and using the first stone tools.
Earliest Homo
The reconstructed Ledi Geraru mandible (top left), compared with Homo naledi (top right), A. deyiremeda (bottom left), and the Uraha early Homo mandible from Malawi (bottom right). Jaws are scaled to roughly the same length from the front to back teeth; the Uraha mandible does not have an erupted third molar whereas the others do and are fully adult.
Just as Sonia Harmand and colleagues pushed back the origins of technology, Brian Villmoare et al. pushed back the origins of the genus Homo, with a 2.7 million year old mandible from Ledi Geraru in Ethiopia. This fossil is only a few hundred thousand years younger than Australopithecus afarensis fossils from the nearby site of Hadar. But the overall anatomy of the Ledi Geraru jaw is quite distinct from A. afarensis, and is much more similar to later Homo fossils (see image above). Hopefully 2016 will reveal other parts of the skeleton of whatever species this jaw belongs to, which will be critical in helping explain how and why our ancestors diverged from the australopithecines. (note that we don’t yet have a date for Homo naledi – maybe these will turn out to be older?)
Early and later Homo
Left: modified figures 2-3 from Maddux et al. (2015). Right: modified figures 7 & 13 from Ward et al. (2015). Note that in the right plot, ER 5881 femur head diameter is smaller than all other Homo except BSN 49/P27.
The earlier hominin fossil record wasn’t the only part to be shaken up. A small molar (KNM-ER 51261) and a set of associated hip bones (KNM-ER 5881) extended the lower range of size variation in Middle and Early (respectively) Pleistocene Homo. It remains to be seen whether this is due to intraspecific variation, for example sex differences, or taxonomic diversity; my money would be on the former.
Left: Penghu 1 hemi-mandible (Chang et al. 2015: Fig. 3), viewed from the outside (top) and inside (bottom). Right: Manot 1 partial cranium (Hershkovitz et al. 2015: Fig. 2), viewed from the left (top) and back (bottom).
At the later end of the fossil human spectrum, researchers also announced an archaic looking mandible dredged up from the Taiwan Straits, and a more modern-looking brain case from Israel. The Penghu 1 mandible is likely under 200,000 years old, and suggests a late survival of archaic-looking humans in East Asia. Maybe this is a fossil Denisovan, who knows? What other human fossils are waiting to be discovered from murky depths?
The Manot 1 calvaria looks very similar to Upper Paleolithic European remains, but is about 20,000 years older. At the ESHE meetings, Israel Hershkovitz actually said the brain case compares well with the Shanidar Neandertals. So wait, is it modern or archaic? As is usually the case, with more fossils come more questions.
Crazy dinosaurs
Yi qi was bringing Skeksi back, and its upper limb had a wing-like shape not seen in any other dinosaur, bird or pterosaur. There were a number of other interesting non-human fossil announcements in 2015 (see here and here), proving yet again that evolution is far more creative than your favorite monster movie makers.
What a year – new species, new tool industries, new ranges of variation! 2015 was a great year to be a paleoanthropologist, and I’ll bet 2016 has just as much excitement in store.
References (in order of appearance)
Haile-Selassie, Y., Gibert, L., Melillo, S., Ryan, T., Alene, M., Deino, A., Levin, N., Scott, G., & Saylor, B. (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity Nature, 521 (7553), 483-488 DOI: 10.1038/nature14448
Harmand, S., Lewis, J., Feibel, C., Lepre, C., Prat, S., Lenoble, A., Boës, X., Quinn, R., Brenet, M., Arroyo, A., Taylor, N., Clément, S., Daver, G., Brugal, J., Leakey, L., Mortlock, R., Wright, J., Lokorodi, S., Kirwa, C., Kent, D., & Roche, H. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521 (7552), 310-315. DOI: 10.1038/nature14464
McPherron, S., Alemseged, Z., Marean, C., Wynn, J., Reed, D., Geraads, D., Bobe, R., & Béarat, H. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature, 466 (7308), 857-860. DOI: 10.1038/nature09248
Thompson, J., McPherron, S., Bobe, R., Reed, D., Barr, W., Wynn, J., Marean, C., Geraads, D., & Alemseged, Z. (2015). Taphonomy of fossils from the hominin-bearing deposits at Dikika, Ethiopia Journal of Human Evolution, 86, 112-135 DOI: 10.1016/j.jhevol.2015.06.013
Villmoare, B., Kimbel, W., Seyoum, C., Campisano, C., DiMaggio, E., Rowan, J., Braun, D., Arrowsmith, J., & Reed, K. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia Science, 347 (6228), 1352-1355 DOI: 10.1126/science.aaa1343
Maddux, S., Ward, C., Brown, F., Plavcan, J., & Manthi, F. (2015). A 750,000 year old hominin molar from the site of Nadung’a, West Turkana, Kenya Journal of Human Evolution, 80, 179-183 DOI: 10.1016/j.jhevol.2014.11.004
Ward, C., Feibel, C., Hammond, A., Leakey, L., Moffett, E., Plavcan, J., Skinner, M., Spoor, F., & Leakey, M. (2015). Associated ilium and femur from Koobi Fora, Kenya, and postcranial diversity in early Homo Journal of Human Evolution, 81, 48-67 DOI: 10.1016/j.jhevol.2015.01.005
Chang, C., Kaifu, Y., Takai, M., Kono, R., Grün, R., Matsu’ura, S., Kinsley, L., & Lin, L. (2015). The first archaic Homo from Taiwan Nature Communications, 6 DOI: 10.1038/ncomms7037
Hershkovitz, I., Marder, O., Ayalon, A., Bar-Matthews, M., Yasur, G., Boaretto, E., Caracuta, V., Alex, B., Frumkin, A., Goder-Goldberger, M., Gunz, P., Holloway, R., Latimer, B., Lavi, R., Matthews, A., Slon, V., Mayer, D., Berna, F., Bar-Oz, G., Yeshurun, R., May, H., Hans, M., Weber, G., & Barzilai, O. (2015). Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans Nature, 520 (7546), 216-219 DOI: 10.1038/nature14134
I went to a cafe today to eat breakfast and get some work done. Write, write, write. It’s important to be properly nourished to ensure maximal productivity.
The Ron Swanson diet.
But I was aghast to behold the food they placed before me:
What on earth is this?
First of all, this is not a sufficient amount of bacon.
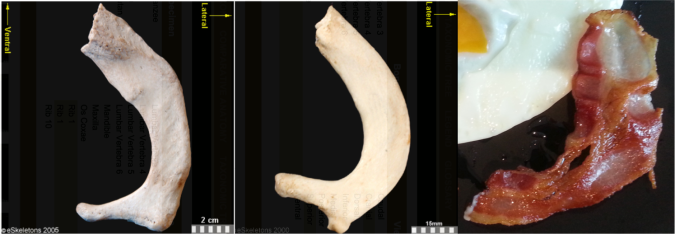
Secondably, this bacon is a spitting image of a first rib:
First ribs from the right side of the body, viewed from the top. From left to right: Human, chimpanzee, bacon. First two images from eSkeletons.org.
At the top of the ribcage, just beneath the clavicle and subclavian artery and vein, the first rib is much shorter and flatter than the rest of the ribs. As Jess Beck at Bone Broke points out, “The first and second rib give something of an awkward ‘slow song at a middle-school dance’ kind of a hug, while the lower ribs provide a more comfortable and self-assured embrace.” I mean, just lookit how sheepishly the bacon dances with the eggs in the first picture, it has ‘middle-school dance’ written all over it.
But the bacon is not totally identical to the human and chimpanzee counterparts. It’s missing their anteromedially sweeping arc, and the distal portion reaching out to the egg is fairly straight. This suggests we’re probably missing much of the original distal end. Posteriorly or dorsally (toward the bottom in the pic), it also appears to be missing much of the lateral portion including the vertebral facet. In this regard, this bacon rib looks a lot like the first rib of Homo naledi:
Full stack of ribs. Left to right: Human, bacon, Homo naledi, Dmanisi Homo erectus, Australopithecus sediba (x2), Australopithecus afarensis specimen “Lucy,” Ardipithecus ramidus, and chimpanzee. Images not to scale except Lucy and Ardi. Image credits given below.
It is hard to make good homologous comparisons among these fossils and bacon, since so many are so incomplete. But it looks like the hominins are relatively longer (front to back, or dorsoventrally) compared to the chimpanzee. That is, oriented along the rib “neck,” the ventral/distal end projects far more medially beyond the proximal vertebral facet in the chimp, while in the hominins the two ends are more flush. Ardi is really incomplete and so very hard to orient, but it may be more like the chimp (I think it needs to be rotated to the right more, to make the lateral edge more vertical like all the other specimens).
It will be interesting to see what my colleagues working on the Homo naledi thorax have to say about rib shapes and their functional importance, hopefully not too long from now.
Anyway, I really wish I had more bacon.
Fossil rib sources Dmanisi Homo erectus: Lordkipanidze D, Jashashvili T, Vekua A, Ponce de León MS, Zollikofer CP, Rightmire GP, Pontzer H, Ferring R, Oms O, Tappen M, Bukhsianidze M, Agusti J, Kahlke R, Kiladze G, Martinez-Navarro B, Mouskhelishvili A, Nioradze M, & Rook L (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449 (7160), 305-10 PMID: 17882214
Australopithecus sediba: Schmid P, Churchill SE, Nalla S, Weissen E, Carlson KJ, de Ruiter DJ, & Berger LR (2013). Mosaic morphology in the thorax of Australopithecus sediba. Science, 340 (6129) PMID: 23580537
Australopithecus afarensis and Ardipithecus ramidus: White TD, Asfaw B, Beyene Y, Haile-Selassie Y, Lovejoy CO, Suwa G, & WoldeGabriel G (2009). Ardipithecus ramidus and the paleobiology of early hominids. Science, 326 (5949), 75-86 PMID: 19810190
Last week, I introduced my Human Evolution students to the “robust” australopiths. It was a very delicate time, when we had to have a grown up, mature conversation about adult things. I reminded the students that they’re only human, but they must resist urges that seem so natural. No matter how much they want to, even if their friends are doing it, they must not act on the deep, dark desire to say that “robust” vs. “gracile” Australopithecus differ in their body build.
Don’t do it, Homo naledi. Don’t talk about body size when you mean to talk about jaw and tooth size. Illustration by Flos Vingerhoets.
Every semester, students (who don’t read and/or pay attention to lecture) think that the difference between these two groups has to do with the species’ body sizes. This is a misconception that has reached the highest echelons of reference:
Apple and Google, at least one person here is not citing their source: F-. Also, is no one else surprised that this term is allegedly specific to anthropology?
No. In the case of australopiths, “gracile” and “robust” refer to the relative size of the jaws, teeth and chewing muscles (all contributing to the “masticatory apparatus”). Traditionally, graciles include the ≥2 million year old Australopithecus afarensis and africanus, and robusts include the later A. boisei and robustus. The discovery of an A. aethiopicus cranium (Walker et al. 1986) somewhat blurred the lines between the two groups but it is usually included with the robusts (who are often collectively called Paranthropus). John Fleagle’s classic textbook (1999) illustrates the gracile-robust dichotomy very nicely:
Comparison of gracile (left) and robust (right) craniodental traits. From Fleagle, 1999.
So to recap: Jaws and teeth, people! To the best of my knowledge, there’s little or no evidence that the various australopithecines differed appreciably in body size (McHenry and Coffing, 2000), stoutness, or muscularity. Although the OH 80 partial skeleton, attributed to Australopithecus boisei based on tooth size and proportions, includes a humerus with very thick cortical bone and a radius with a crazy big insertion for the biceps muscle—it was a very large and muscular A. boisei (Domínguez-Rodrigo et al., 2013). Nevertheless, gracile and robust australopithecine species differ most notably in their jaws and teeth, not bodies. Maybe this is why Liz Lemon was so confused about the term “robust”?
Today, these are somewhat antiquated terms. Back when the only early hominins known to science were the species listed above, it was easy to make a distinction. However, as the fossil record has expanded of late, the gracile-robust dichotomy becomes blurry. Australopithecus garhi (Asfaw et al., 1999) has overall tooth proportions comparable to graciles, but absolute tooth sizes and sagittal cresting like robusts. The recently described Australopithecus deyiremeda has tooth sizes and proportions like graciles but lower jaws that are very thick, like those of robust australopiths (Haile-Selassie et al., 2015).
So in light of all the confusion and blurring distinctions, maybe it’s time to scrap “gracile” vs. “robust”?
References Asfaw B, White T, Lovejoy O, Latimer B, Simpson S, & Suwa G (1999). Australopithecus garhi: a new species of early hominid from Ethiopia. Science (New York, N.Y.), 284 (5414), 629-35 PMID: 10213683
Domínguez-Rodrigo, M., Pickering, T., Baquedano, E., Mabulla, A., Mark, D., Musiba, C., Bunn, H., Uribelarrea, D., Smith, V., Diez-Martin, F., Pérez-González, A., Sánchez, P., Santonja, M., Barboni, D., Gidna, A., Ashley, G., Yravedra, J., Heaton, J., & Arriaza, M. (2013). First Partial Skeleton of a 1.34-Million-Year-Old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania PLoS ONE, 8 (12) DOI: 10.1371/journal.pone.0080347
Haile-Selassie Y, Gibert L, Melillo SM, Ryan TM, Alene M, Deino A, Levin NE, Scott G, & Saylor BZ (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity. Nature, 521 (7553), 483-8 PMID: 26017448
Walker, A., Leakey, R., Harris, J., & Brown, F. (1986). 2.5-Myr Australopithecus boisei from west of Lake Turkana, Kenya Nature, 322 (6079), 517-522 DOI: 10.1038/322517a0
It will be lots of work to prep my Human Evolution course for the Fall. This past year has seen many major fossil discoveries, and adding to the list is the newly described species Australopithecus deyiremeda (Haile-Selassie et al., 2015). The fossils come from newly announced sites in Ethiopia (here it is on a map!), dating to around 3.4 million years ago. These new fossils are contemporaneous with Australopithecus afarensis, fossils attributed to Kenyanthropus platyops, and whatever the hell the Burtele foot belongs to.
The main specimens are a fairly complete half of a maxilla (upper jaw) and two decent mandibles (lower jaw bones). These fossils do not belong to the same individual (despite all the media pictures of the upper and lower jaws together). One of the most distinctive features of these fossils is how thick, both in absolute and relative terms, the mandibles are, especially given how short they are. What sticks out to me though, is that the upper jaw looks like it might have still had some growing to do. Why on earth would I think so? (The following is based off pictures from the publications, so I could be wrong!)
Extended Figure 1a from the paper, the type specimnen BRT-VP-3/1 maxilla viewed from the left side. I’ve added the M2 label for your reading pleasure.
The holotype maxilla (BRT-VP-3/1) is described as coming from a “young adult” in the Supplementary Information. However, it looks like the second molar tooth (M2) is not quite fully erupted and in occlusion, although this could be due to the natural arc of the tooth row. There is no visible wear on the tooth in the pictures, and indeed the Supplementary Information says the tooth is unworn. This means that the tooth is only recently emerged, and may not have passed the gum line, and therefore hasn’t seen much/any use yet. Authors note in the Supplementary Information that there is no M3 (a.k.a. “wisdom tooth”) wear facet on the back of M2 , meaning the last tooth hadn’t yet emerged yet either. So, this all points to a non-adult age by tooth eruption standards.
Extended Figure 1d from the paper. Same fossil, but from the bottom; pretend you’re a dentist peering into its mouth. Back is to the bottom.
In addition, the M2 roots don’t look fully formed. This is especially apparent in Extended Figure 1h, a CT section through the teeth:
Left side: Extended Figure 1h from the paper. From left to right, the teeth are P3, P4, M1, and M2. For comparison, to the right are Demirjian tooth development stages, modified from Table 2 of Kuykendall, 1996. Also compare the M2 roots with completed roots of the M1.
In many human populations, this stage of M2 development is reached (on average) between 11-13 years (Liversidge et al., 2006). In the wild Taï Forest chimpanzee sample, two individuals with M2 root completely formed (Stage H) are 10 and 11 years old (Smith et al., 2010). These apes would not be fully mature and their facial dimensions would likely have increased had they reached adulthood (Zihlman et al., 2007).
So what this suggests to me is that this maxilla may not accurately represent adult anatomy in this newly described species. In humans, the face continues to grow downwards from adolescence into adulthood, and in apes the face continues growing both forward and downward. In the differential diagnosis of A. deyiremeda, Haile-Selassie and team state, in layman’s terms, that the cheeks are positioned more toward the front than in A. afarensis, and that the front of the face doesn’t stick out as much as in A. garhi. If this specimen was not fully grown, it is likely that the true adult anatomy would have had a face that sticks out more and has less forward-positioned cheeks than in this specimen.
But, this is all speculative, and I’d like to reiterate that these observations of dental development are based only on the published pictures. Just a thought!
Matt Skinner and colleagues report in today’s Science an analysis of trabecular bone structure in the hand bones of humans, fossil hominins and living apes. Trabecular bone, the sponge-like network of bony lattices on the insides of many of your bones, adapts during life to better withstand the directions and amounts of force it experiences. This is a pretty great property of the skeleton: bone is organized in a way that helps withstand usual forces, and the spongy organization of trabeculae also keeps bones fairly lightweight. Win-win.
An X-ray of my foot. The individual foot bones are filled with narrow spicules (=trabeculae) of bone. Very often they have a directed, or non-random, orientation: in the calcaneus, for instance, they are oriented mostly from the heel to the ankle joint.
This adaptive nature of trabecular bone also means that we can learn a lot about how animals lived in the past when all they’ve left behind are scattered fossils. In the present case, Skinner and colleagues tested whether tool use leaves a ‘trabecular signature’ in hand bones, looking then for whether fossil hominins fit this signature. Their study design is beautifully simple but profoundly insightful: First, they compared humans and apes to see if the internal structure of their hand bones can be distinguished. Second, they tested whether these differences accord with theoretical predictions based on how these animals use their hands (humans manipulate objects, apes use hands for walking and climbing). Third, they determined whether fossil hand bones look more like either group.
Comparison of first metacarpals (the thumb bone in your palm) between a chimpanzee (left), three australopithecines, and a human (right). In each, the palm side is to the left and the wrist end of the bone (proximal) is down. Image by Tracy Kivell, and found here.
Looking at the image above, it’s difficult to spot trabecular differences between the specimens with the naked eye. But computer software can easily measure the density and distribution of trabecular bone from CT scans. With these tools, researchers found key differences between humans and apes consistent with the different ways they use their hands. Neandertals (humans in the past 100 thousand years or so) showed the human pattern, not unexpected since their bones look like ours and they used their hands to make tools and manipulate objects like we do.
What’s more interesting, though, is that the australopithecines, dating to between 1.8-3.0 million years ago, also show the human pattern. This is an important finding since the external anatomy of Australopithecus hand bones shows a mixture of human- and ape-like features, with unclear implications for how they used their hands. Their trabecular architecture, reflecting the forces their hands experienced in life, is consistent with tool use.
This is a very significant finding.Australopithecus africanus fossils from Sterkfontein aren’t associated with any stone tools; bone tools are known from Swartkrans, though it is unclear whether Australopithecus robustus or Early Homo from the site made/used these. In addition, in 2010 McPherron and colleagues reported on a possibly cut-marked animal bone from the 3.4 million year old site of Dikika in Ethiopia, where Australopithecus afarensis fossils but no tools are found. Skinner and colleagues’ results show that at the very least, South African Australopithecus species were using their hands like tool-makers and -users do.
This raises many fascinating questions – were australopithecines using stone tools, but we haven’t found them? Were they using tools made of other materials? What do the insides of Australopithecus afarensis metacarpals look like? What I like about this study is that it presents both compelling results, and raises further (testable) questions about both the nature of the earliest tools and our ability to detect their use from fossils.
Earlier this year I suggested a classroom activity in which students can scrutinize the evidence used to argue that the >5 million year old (mya) Ardipithecus kadabba was bipedal. To recap: Ar. kadabba is represented by some teeth, a broken lower jaw, and some fragmentary postcrania. The main piece of evidence that it is a human ancestor and not just any old ape is from a single toe bone, and the orientation of its proximal joint. In Ar. kadabba and animals that hyperdorxiflex their toes (i.e., humans and other bipeds when walking), this joint faces upward, whereas it points backward or even downward in apes. This “dorsal canting” of the proximal toe joint has also been used as evidence that the 4.4 mya Ardipithecus ramidus and 3.5 mya owner of the mystery foot from Burtele are bipedal hominins. A question remains, though – does this anatomy really distinguish locomotor groups such as bipeds from quadrupeds?
STUDENT SCIENTISTS TO THE RESCUE! Use ImageJ to measure the canting angle between the proximal joint and plantar surface, as I’ve done on this Japanese macaque monkey (they are not bipedal). Proximal to the right, distal to the left Note I changed the measured angle from the March post.
I sicked my students in Ant 364 (Human Evolutionary Developmental Biology) here at NU on this task. I had students look at only 11 modern primates from the awesome KUPRI database. Most groups are only represented by 1 (Homo sapiens, Hylobates lar and Macaca fuscata) or two (Pongo species and Gorilla gorilla) specimens, all adults. For chimpanzees (Pan troglodytes) there is one infant and four adults. The database has more individuals, and it would be better to include more specimens to get better ideas of species’ ranges of variation, but this is a good training sample for a class assignment. The fossil group includes one Ardipithecus ramidus,one Ar.kadabba, one Australopithecus afarensis, and the PPP4 of the mystery foot from Burtele. The human and all fossils except Ar. kadabba are based off of lateral photographs and not CT scans like for the living primates, meaning there may be some error in their measurements, but we’ll assume for the assignment this is not a problem. Here are their results:
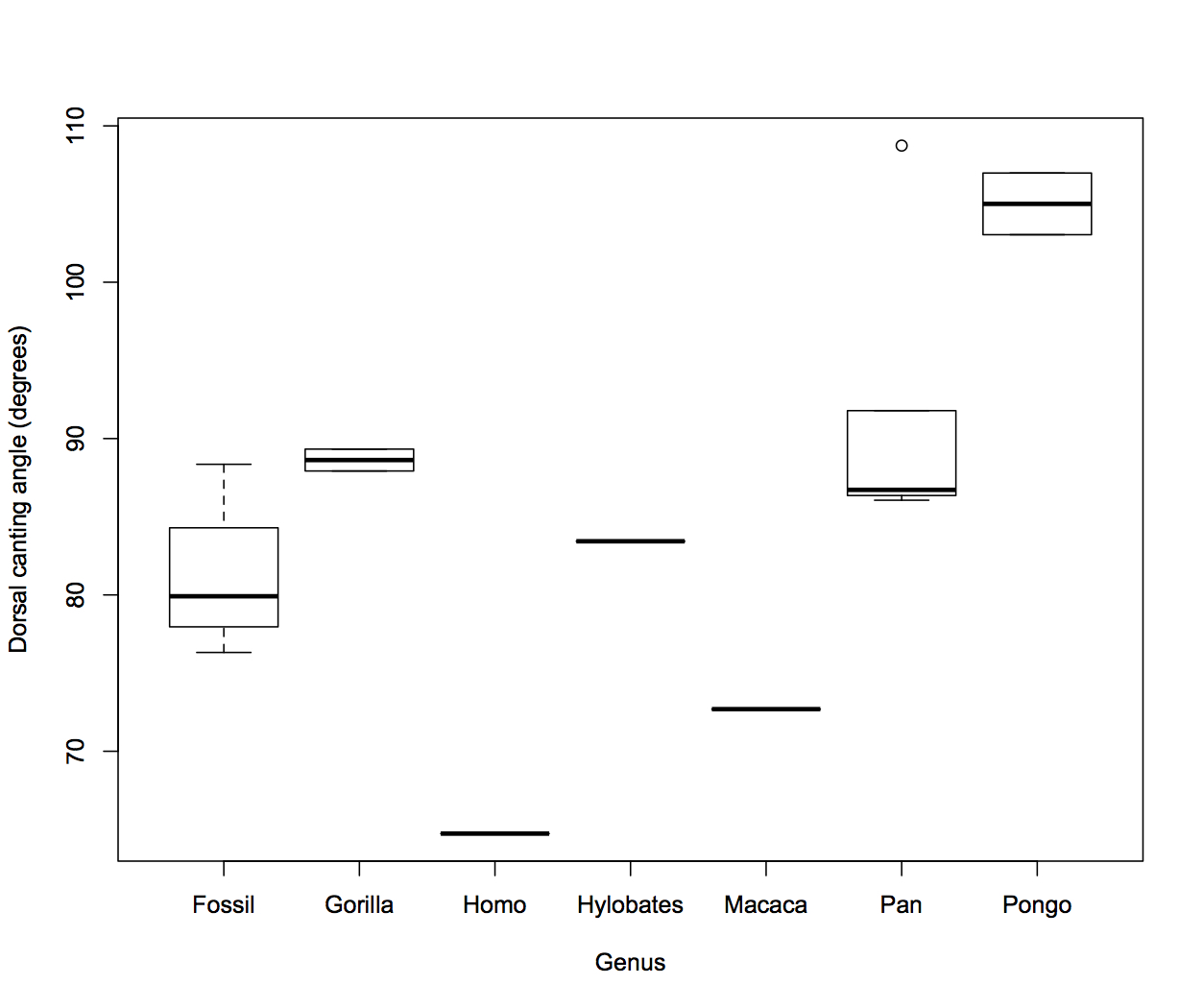
Dorsal canting angle of the fourth proximal pedal phalanx in primates. The lower the angle, the more dorsally canted the proximal joint surface. The “Fossil” group includes specimens attributed to Ardipithecus, Australopithecus and something unknown.
Great apes have fairly high angles, meaning generally not dorsally canted proximal joint surfaces. The two gorillas fall right in the adult chimpanzee (adult) range of variation, while chimp infant and orangutans have much higher angles (≥90º means they’re actually angled downward or plantarly). The gibbon (Hylobates) is slightly lower than the chimpanzee range. The macaque has an even more dorsally canted joint, and the human even more so. The fossils, except the measurement for Ar. ramidus (see note above), have lower angles than living apes, but higher than the human and the monkey. If dorsal canting really is really a bony adaptation to forces experienced during life, then the fossil angles suggest these animals’ toes were dorsiflexed more so than living great apes (but not as much as the single monkey and human).
This lab helps students become familiar with CT data, the fossil record, taking measurements (students also measure maximum length of the toe bones and look at the relationship between length and canting), analyzing data, and hypothesis testing. You can also have fun exploring inter-observer error by comparing students’ measurements.
Last week I was exploring central England with the brilliant Jess Beck, an archaeology PhD student at the University of Michigan. Both of us avid (nay, rabid) connoisseurs of everything skeletal, we espied the likes of a specific human bone in the scenic landscape of the the Cotswolds. Check out JB’s blog, Bone Broke, for her take on this geographical/geological/skeletal formation (as well as for lots of killer osteology and bioarchaeology tips and tricks). Do it now! NOW!
After you’ve checked out her site, behold this sight – what bone is lurking in the landscape?
As with Rorschach inkblots, probably lots of bones could be seen in this image. But what Jess & I saw was a hamate, the greener hue hewn into the hills, whose sizable hamulus runs from the bottom right to join the rest of the carpal around the center of the image.
The hamulus of the hamate is an attachment point for the flexor retinaculum, the band of fascia stretching across your wrist to hold your extrinsic digital flexor muscles (or rather, their tendons) in place; you could think of it as the bridge covering the carpal tunnel. Now, comparing the grassy hamulus with an actual human one, you’ll spot two important differences: first, the grassy one isn’t blunt like the humans’, but ends in a long point. Oops! Just pretend it’s rounded off. Second, the grassy hamulus is huge relative to the overall size of the bone (or valley) compared with the human form. The size of the hamulus partially reflects the size of the carpal tunnel: chimpanzees, with powerful wrists and forearms, have long hamuli.
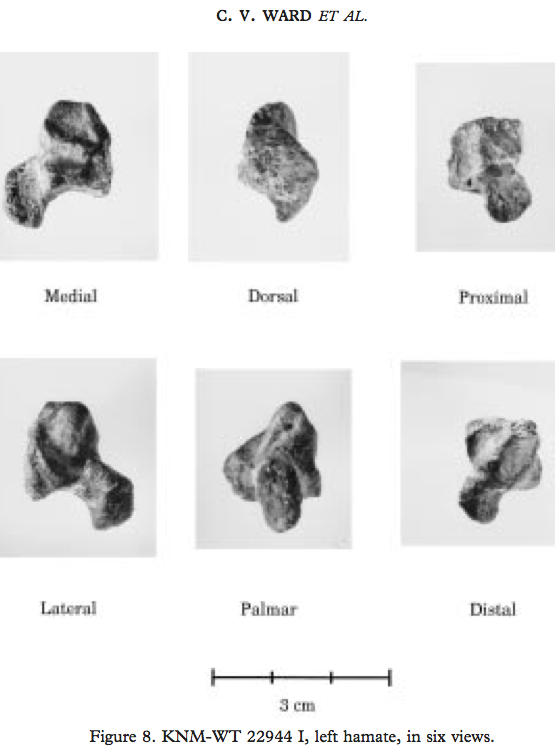
A huge nerd, I didn’t just see any hamate in this Cotswold vale. I also immediately thought of KNM-WT 22944, an Australopithecus afarensis hamate from the 3.5 million year old site of South Turkwel in Kenya (Ward et al., 1997):
From Ward et al., 1999. Sorry it’s not in the same orientation as the above image. Hamulus is the projection pointing to the bottom left corner of the “medial” image.
An absolutely and relatively massive hamulus in WT 22944 suggests whoever this bone belonged to had some powerful gripping capabilities, while a geologically younger A. afarensis hamate from Hadar (AL 333-50) had a smaller, more human-like hamulus. Maybe (some) A. afarensis were still using their arms a lot for tree-climbing, in spite of being more than capable bipeds (I’ve talked about this before here)….