A week and a half ago, Kibii and colleagues (2011) published reconstructions and re-analyses of two hips belonging to the 1.98 million-year old Australopithecus sediba. As with many fossil discoveries, these additions to the fossil record raise more questions than they answer. Unless the question was, “did A. sediba have a pelvis?” It did. Here’s a good summary from the paper itself:
Thus, Au. sediba is australopith-like in having a long superior pubic ramus and an anteriorly positioned and indistinctly developed iliac pillar…[and] Homo-like in having vertically oriented and sigmoid shaped iliac blades, more robust ilia, and a narrow tuberoacetabular sulcus…and the pubic body is upwardly rotated as in Homo. (p. 1410, emphases mine)
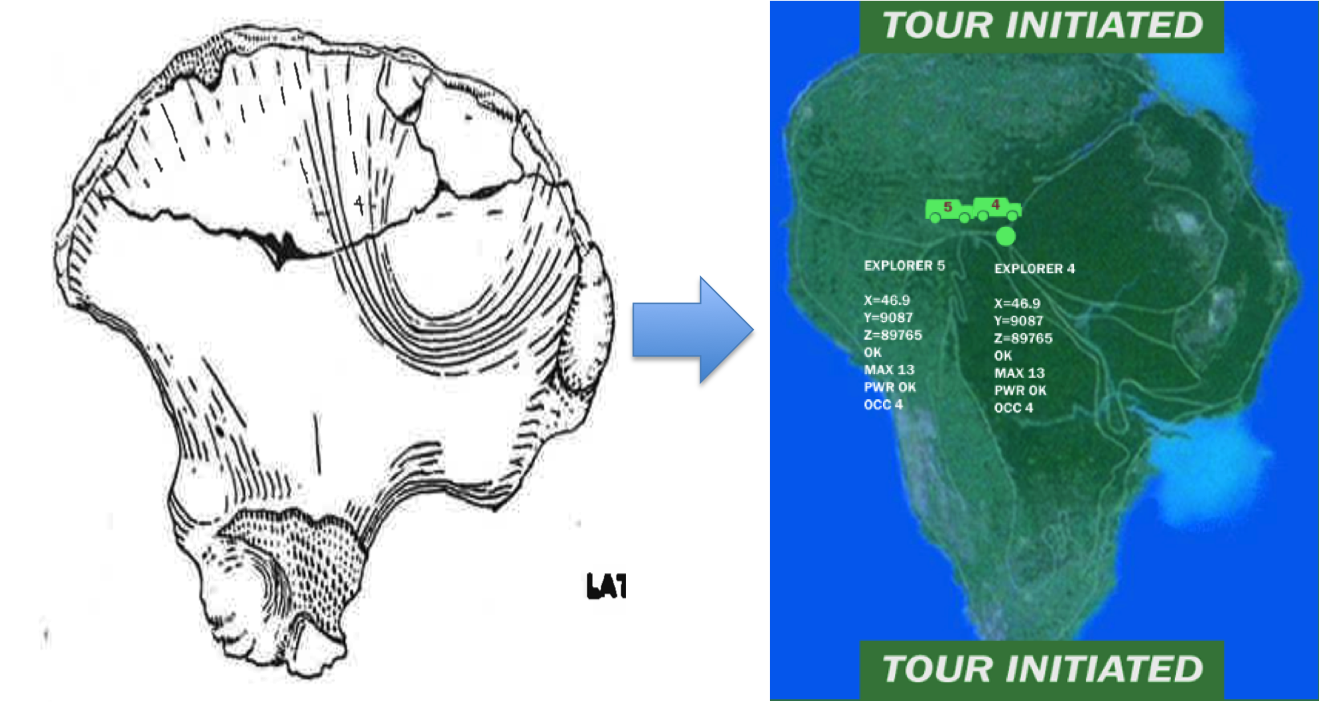
So far as I can tell, the main way the hips are ‘advanced’ toward a more human-like condition is that the iliac blades are more upright and sweep forward more than in earlier known hominid hips. Here’s the figure 2 from the paper (more sweet pics of the fossils are available here). NB that in both A. sediba hips much of the upper portions of the iliac blades are missing (reconstructed in white; this region is missing in lots of fossils), so it’s possible they were more flaring like the australopith in the center photo.

The authors’ bottom-line, take-home point is that the A. sediba pelvis has features traditionally associated with large-brained Homo – but belonged to a small-brained species (based solely on the ~430 cc MH1 endocast). They argue that this means that many of these unique pelvic features did not evolve in the context of birthing large-brained babies, as has often been thought. They state that these features are thus “most parsimoniously attributed to altered biomechanical demands on the pelvis in locomotion,” and suggest that this hypothetical locomotion was mostly bipedalism but with a good degree of climbing. Maybe, maybe not. This interpretation is consistent with the analysis of the A. sediba foot/ankle (Zipfel et al. 2011).
The weird mix of ancient (australopith-like) and newer (Homo-like) pelvic features in A. sediba really raises the question of how australopithecines moved around. More intriguing is that the A. sediba pelvis has different Homo-like features than the ~1 million year old Busidima pelvis (Simpson et al. 2008), which has been attributed to Homo erectus (largely in aspects of the iliac blades). This raises the question of whether A. sediba is really pertinent to the origins of the genus Homo, and whether the Busidima pelvis belongs to Homo erectus or a late-surviving robust australopithecus (e.g. boisei, Ruff 2010).
Also interesting is that the subpubic angle (in the pic above, the upside-down “V” created by the pubic bones just above the red labels) is pretty low in MH2. This is curious because modern human males and females differ in how large this angle is – females tend to have a large angle which contributes to an enlarged birth canal, whereas males have a low angle like MH2. But MH2 is considered female based on skeletal and dental size. This raises the additional questions of whether human-like sexual dimorphism had not evolved in hominids prior to 1.9 million years ago, and whether the sex of MH2 was accurately described.
Finally, though the authors did a great job comparing this pelvis with those from other hominids, I think a major, more comprehensive comparative review of hominid pelves is in order. How does the older A. afarensis hip from Woranso (Haile-Selassie et al. 2010) inform australopithecine pelvic evolution? What about the possibly-contemporary-maybe-later hip from the nearby site of Drimolen (Gommery et al. 2002)? Given the subadult status of the MH1 individual, it would be interesting to compare with the WT 15000 Homo erectus fossils, or A. africanus subadults from Makapansgat, to examine the evolution of pelvic growth.

Lots of interesting questions arise from these fascinating new fossils. “The more you know,” right?
References
Gommery, D. (2002). Description d’un bassin fragmentaire de Paranthropus robustus du site Plio-Pléistocène de Drimolen (Afrique du Sud)A fragmentary pelvis of Paranthropus robustus of the Plio-Pleistocene site of Drimolen (Republic of South Africa) Geobios, 35 (2), 265-281 DOI: 10.1016/S0016-6995(02)00022-0
Haile-Selassie Y, Latimer BM, Alene M, Deino AL, Gibert L, Melillo SM, Saylor BZ, Scott GR, & Lovejoy CO (2010). An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia. Proceedings of the National Academy of Sciences of the United States of America, 107 (27), 12121-6 PMID: 20566837
Kibii, J., Churchill, S., Schmid, P., Carlson, K., Reed, N., de Ruiter, D., & Berger, L. (2011). A Partial Pelvis of Australopithecus sediba Science, 333 (6048), 1407-1411 DOI: 10.1126/science.1202521
Ruff, C. (2010). Body size and body shape in early hominins – implications of the Gona Pelvis Journal of Human Evolution, 58 (2), 166-178 DOI: 10.1016/j.jhevol.2009.10.003
Simpson, S., Quade, J., Levin, N., Butler, R., Dupont-Nivet, G., Everett, M., & Semaw, S. (2008). A Female Homo erectus Pelvis from Gona, Ethiopia Science, 322 (5904), 1089-1092 DOI: 10.1126/science.1163592
Zipfel, B., DeSilva, J., Kidd, R., Carlson, K., Churchill, S., & Berger, L. (2011). The Foot and Ankle of Australopithecus sediba Science, 333 (6048), 1417-1420 DOI: 10.1126/science.1202703