Two reports came out last week in the journal Nature, re-dating some European human fossils to before 40 thousand years ago (kya), a few thousand years older than previous evidence for modern-looking people in the region. The media have been reporting these studies as revealing “the first Europeans,” but of course we all know that the first Europeans were the badass hominids, my favorites, from the 1.8 million year old site of Dmanisi.
 |
| KC4 maxilla (Higham et al. 2011) |
From Kent’s Cavern (United Kingdom) is a partial maxilla, now dated to 44 – 41 kya* (Higham et al. 2011; but see below). The jaw fragment with highly worn teeth was found just above some Aurignacian-like (Upper Paleolithic) blades in 1927. [NB below these 2 blades were 2 other blades of a tool “complex…tentatively associated with Neandertals] The laughable amount of bone makes it rather impossible to say whether the fossil represents a Neandertal or more modern-looking human. The authors examined what little of the morphology was left and concluded that the fossil shared the most similarities with recent humans but only a few with Neandertals. A more rigorous analysis of what this mix of traits means would have been nice (i.e. why would an individual have derived traits of both ‘modern’ humans and Neandertals?). The researchers tried to extract DNA for analysis, but apparently organic remains were too poorly preserved for a good analysis. Bummer.
 |
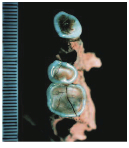
| Cavallo B and C (Benazzi et al. 2011) |
A similar older-than-we-thought story is reported by Stefano Benazzi and buddies, who reanalyzed teeth from the Italian site of Grotta del Cavallo (left). After the site was excavated in 1967, the teeth were attributed to Neandertals and the lithics classified as “Uluzzian.” I would be a dirty liar if I said knew anything about the “Uluzzian” industry (try this other site which may be more informative), but apparently it’s seen as transitional between the Neandertal-associated Mousterian and ‘more advanced’ Upper Paleolithic toolkits. So this assemblage could be used to argue that Neandertals were smart enough to upgrade to a sexier stone tool industry shortly before their anatomy (but not their genes!) disappeared. BUT! also like in Kent’s Cavern paper, Benazzi and colleagues examined what little morphology is preserved in the fossil teeth, and (re)assigned them to modern-looking humans. The authors provided nice qualitative and quantitative arguments for the human status of the teeth, though again I have to raise caution that these are only teeth and we have no idea what the rest of the skeleton would have looked like. Researchers also analyzed shells associated with the now-human teeth and dated the site to around 44 kya, making them the oldest probably-human remains in Europe.
Now, according to conventional wisdom, the Neandertals were dullards who made and used the Mousterian stone tool industry. The Mousterian was nice and all, but not nearly as wicked-pisser as the smart and sassy modern-looking humans’ Aurignacian toolkit. The thing is, though, there really hasn’t been really a lot of evidence unequivocally linking modern-looking fossils with Aurignacian artifacts. So both of the recent studies in Nature lend support to the idea that maybe modern humans were the sole makers (and users) of an advanced stone tool industry. But it’s important to remember [1] that the blades ‘associated’ with the Kent’s Cavern jaw can’t really be definitively attributed to a stone tool industry; [2] the blades were deeper in the cave than the jaw, and so may actually be appreciably older than the jaw; and [3] while the teeth from Kent’s Cavern and Cavallo do look most comparable to those belonging to ‘anatomically’ modern humans, we don’t know what the skulls or skeletons containing the teeth looked like. All that said, it’s neat to see the possible appearance of certain anatomy and technology in Europe thousands of years earlier than previously thought. It also re-raises the issue of the degree to which modern-looking humans and Neandertals overlapped in space and time, and what these interactions would have been like (I’d guess terrifying, hilarious and/or sexy).
 |
| KC4 originally (Higham et al. 2011) |
A fun lesson also comes from the Kent’s Cavern study. Higham and colleagues report that the KC4 human maxilla was excavated in 1927 and analyzed by Sir Arthur Keith, a well-trained anatomist and paleoanthropologist famous in his day. Keith described the fossil as containing a canine, second premolar and first molar (right, compare with above). Higham and colleagues, however, noticed that what Keith described as a second premolar was in fact a first premolar. The teeth are so worn they mostly lack information about their form and features, so this mistake probably didn’t really mislead anatomists in any way. Still, it goes to show that even experts like Keith can make mistakes or overlook the things they know best, and this is not the first time I’ve seen this happen with fossils. So always (politely) question those giants whose shoulders you’re standing upon.
*Update (07 Nov): John Hawks points out that the KC4 maxilla itself was not used to obtain the older radiocarbon age estimate. In 1989 the maxilla was directly dated to around 30 kya, over 10 ky younger than the new estimates. Higham and friends thought this date was too young, based on methodological grounds. An attempt to redate the KC4 maxilla based on one of the teeth yielded too little collagen (organic material) to produce a date. Bummer again! The new estimate is instead based on radiocarbon dates obtained from animal bones that were excavated from above and below the human jaw. So this ‘redating’ of the Kent’s Cavern maxilla is very interesting, but should be taken with a grain of salt.
![]()
See for yourself
Benazzi, S., Douka, K., Fornai, C., Bauer, C., Kullmer, O., Svoboda, J., Pap, I., Mallegni, F., Bayle, P., Coquerelle, M., Condemi, S., Ronchitelli, A., Harvati, K., & Weber, G. (2011). Early dispersal of modern humans in Europe and implications for Neanderthal behaviour Nature DOI: 10.1038/nature10617
Higham, T., Compton, T., Stringer, C., Jacobi, R., Shapiro, B., Trinkaus, E., Chandler, B., Gröning, F., Collins, C., Hillson, S., O’Higgins, P., FitzGerald, C., & Fagan, M. (2011). The earliest evidence for anatomically modern humans in northwestern Europe Nature DOI: 10.1038/nature10484
