I went to a cafe today to eat breakfast and get some work done. Write, write, write. It’s important to be properly nourished to ensure maximal productivity.

The Ron Swanson diet.
But I was aghast to behold the food they placed before me:

What on earth is this?
First of all, this is not a sufficient amount of bacon.

Secondably, this bacon is a spitting image of a first rib:
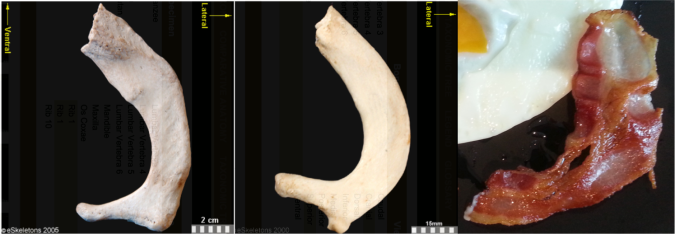
First ribs from the right side of the body, viewed from the top. From left to right: Human, chimpanzee, bacon. First two images from eSkeletons.org.
At the top of the ribcage, just beneath the clavicle and subclavian artery and vein, the first rib is much shorter and flatter than the rest of the ribs. As Jess Beck at Bone Broke points out, “The first and second rib give something of an awkward ‘slow song at a middle-school dance’ kind of a hug, while the lower ribs provide a more comfortable and self-assured embrace.” I mean, just lookit how sheepishly the bacon dances with the eggs in the first picture, it has ‘middle-school dance’ written all over it.
But the bacon is not totally identical to the human and chimpanzee counterparts. It’s missing their anteromedially sweeping arc, and the distal portion reaching out to the egg is fairly straight. This suggests we’re probably missing much of the original distal end. Posteriorly or dorsally (toward the bottom in the pic), it also appears to be missing much of the lateral portion including the vertebral facet. In this regard, this bacon rib looks a lot like the first rib of Homo naledi:

Full stack of ribs. Left to right: Human, bacon, Homo naledi, Dmanisi Homo erectus, Australopithecus sediba (x2), Australopithecus afarensis specimen “Lucy,” Ardipithecus ramidus, and chimpanzee. Images not to scale except Lucy and Ardi. Image credits given below.
It is hard to make good homologous comparisons among these fossils and bacon, since so many are so incomplete. But it looks like the hominins are relatively longer (front to back, or dorsoventrally) compared to the chimpanzee. That is, oriented along the rib “neck,” the ventral/distal end projects far more medially beyond the proximal vertebral facet in the chimp, while in the hominins the two ends are more flush. Ardi is really incomplete and so very hard to orient, but it may be more like the chimp (I think it needs to be rotated to the right more, to make the lateral edge more vertical like all the other specimens).
It will be interesting to see what my colleagues working on the Homo naledi thorax have to say about rib shapes and their functional importance, hopefully not too long from now.
Anyway, I really wish I had more bacon.
Fossil rib sources
![]() Dmanisi Homo erectus: Lordkipanidze D, Jashashvili T, Vekua A, Ponce de León MS, Zollikofer CP, Rightmire GP, Pontzer H, Ferring R, Oms O, Tappen M, Bukhsianidze M, Agusti J, Kahlke R, Kiladze G, Martinez-Navarro B, Mouskhelishvili A, Nioradze M, & Rook L (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449 (7160), 305-10 PMID: 17882214
Dmanisi Homo erectus: Lordkipanidze D, Jashashvili T, Vekua A, Ponce de León MS, Zollikofer CP, Rightmire GP, Pontzer H, Ferring R, Oms O, Tappen M, Bukhsianidze M, Agusti J, Kahlke R, Kiladze G, Martinez-Navarro B, Mouskhelishvili A, Nioradze M, & Rook L (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449 (7160), 305-10 PMID: 17882214
Australopithecus sediba: Schmid P, Churchill SE, Nalla S, Weissen E, Carlson KJ, de Ruiter DJ, & Berger LR (2013). Mosaic morphology in the thorax of Australopithecus sediba. Science, 340 (6129) PMID: 23580537
Homo naledi: Morphosource.
Australopithecus afarensis and Ardipithecus ramidus: White TD, Asfaw B, Beyene Y, Haile-Selassie Y, Lovejoy CO, Suwa G, & WoldeGabriel G (2009). Ardipithecus ramidus and the paleobiology of early hominids. Science, 326 (5949), 75-86 PMID: 19810190