
As mentioned briefly but repeatedly on this blog, my dissertation is about growth of the lower jaw in Australopithecus robustus (right), comparing it with jaw growth in recent humans. This is important because we don’t really know exactly how skeletal-dental (especially skeletal) maturation of our fossil relatives compares with us today. From a developmental perspective, it is also important to know how and when adult form arises during growth, and how these processes vary within and between species.
It’s not easy to examine ontogeny in fossil samples. In a post a few weeks ago I included a drawing of some of the A. robustus juvenile jaws. At the time, I was pointing out variation in dental maturity (which is a nice thing when studying growth), but the picture also reveals a bigger bugbear – variable preservation of features (which is a terrible thing if you’re trying to study growth).

For example, the youngest individual in the fossil sample (right, viewed from above, front is at the top of the picture) includes only the second baby molar tooth, a bit of the bone surrounding the sides and back of the tooth, and a small portion of the ascending ramus. The oldest subadult in the sample (SKW 5), on the other hand, is almost entirely complete. In between these ages, jaws variously preserve different parts. Under these circumstances (i.e. lots of missing data), growth cannot be studied by traditional (namely, multivariate) methods (how I will deal with this is a topic for another day).
So while studying the fossils in South Africa, in order to maximize the number of comparisons I could possibly make, I measured just about every single linear dimension conceivable on these jaws. I thus have a spreadsheet with 300 columns of measurements I could take on each specimen. Most of the cells are empty : (
What’s a boy to do?! In order to compare A. robustus with humans, I need to take the same measurements on a growth series of human jaws, too. But life is short, and if I want to finish this project before I succumb to some sinister signature of senescence, I really can’t take hundreds of measurements on a human sample which is much larger than the fossils. Plus, a lot of the individual measurements are a bit redundant: some of the distances overlap, many of the variables can be taken on the right and the left sides, etc.
If I am to finish collecting data in a reasonable time frame, I need to cull my variables from 300 to however many (a) maximizes the comparisons I can make within the less-complete A. robustus sample, and (b) are not too repetitive. Boo. Plus I have to get these spreadsheets ready to be read and analyzed in the program R, which for whatever reason is always a pain in the ass.
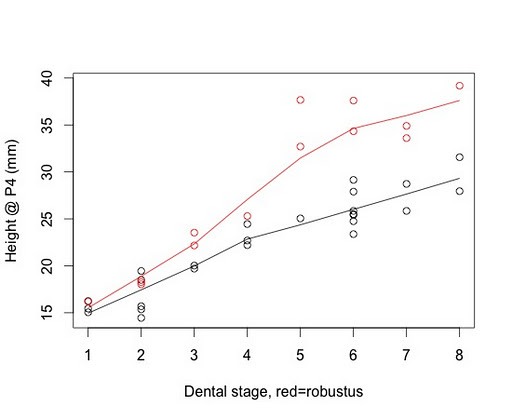
Again, the statistics of the overall comparisons is a topic for another day, and I haven’t had the opportunity yet to write the analytical program(s). But I have looked at some individual traits in A. robustus compared with a subsample of humans. For example, at the left is a plot of changes in height of the jaw at the baby second molar or adult second premolar (which replaces the baby molar). Obviously my human sample is way to small at the moment to make any really meaningful statements about how growth compares between the two species. Note also that these are absolute measures and not size-corrected, and that these are stages of dental eruption rather than chronological ages. But from this preliminary view, the two species are very similar up to around when the first adult molar comes in (“stage 4” here). Thereafter, the A. robustus individuals dramatically increase in size rather fast, whereas humans only slowly increase in size.
Again, this is a very preliminary result, and only for a single measurement. But it is interesting in light of a recent study by Megan Holmes and Christopher Ruff (2011). These researchers compared jaw growth recent humans who differed in the consistency of their diets. They found that kids in the two populations were not too different, but the samples became more different with age to become fairly different as adults. Now, A. robustus seems to have eaten a diet with lots of hard objects (see recent review by Peter Ungar and Matt Spohneimer), but humans’ diet (and technology) really obviates the need for chewing as powerful as seen in A. robustus. So this dietary divergence may well be reflected in the growth difference suggested above, but it may not be the sole factor. PLUS I NEED TO INCREASE MY HUMAN SAMPLE.
Stay tuned for more analyses and results!![]() References to make you smarter and stronger
References to make you smarter and stronger
Holmes, M., & Ruff, C. (2011). Dietary effects on development of the human mandibular corpus American Journal of Physical Anthropology, 145 (4), 615-628 DOI: 10.1002/ajpa.21554
Ungar, P., & Sponheimer, M. (2011) The Diets of Early Hominins. Science 334(6053), 190-193. DOI: 10.1126/science.1207701

{kind=link}