A few weeks ago I posted some early observations I’ve made about mandible growth in Australopithecus robustus compared with humans. My dissertation tests the null hypothesis that overall mandible growth is identical in the two species. This is complicated by the fact that there are many aspects of jaw growth (i.e. lots of variables) and not all fossils preserve the same parts. In these early preparatory stages I’m looking only at the height and width of the jaw at the second baby molar (in kids) and the second permanent premolar that replaces this baby tooth in older individuals, since this is something most of the fossils have. This work will get me ready for the hard comparisons, where the fossils aren’t so kind.
One concern I had in the earlier post was that my human sample was (and still is) fairly small, making comparisons rather tentative. Since then, I have about doubled my human sample (but I still have lots of work to do), so it’s timely to see if my earlier observations have held up. AND THEY DO!

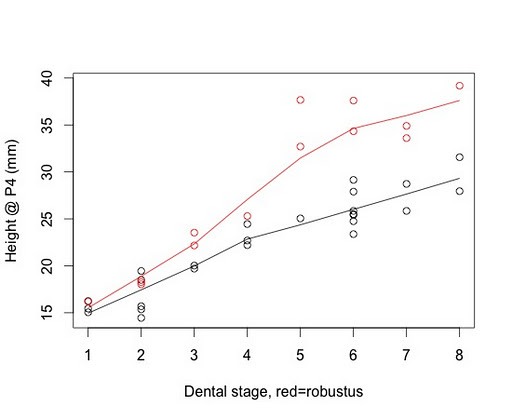
To the right is a plot of jaw height at said tooth position across the growth period, humans being the black circles and A. robustus the thick red ones. Note that measures are standardized, taken relative to the smallest (not necessarily the youngest) individual in each sample. Before, I’d found that the two samples overlapped up to dental stage 4 (when the first permanent tooth comes in). After this point, the A. robustus jaw gets much larger through early adulthood, whereas in humans the height increase isn’t so drastic. With a larger sample, there is a bit more overlap in relative jaw height (especially early on), but the overall result is the same as I found earlier. Neat!

To the left is a similar plot, this time looking at width of the jaw across the growth period (these are also size-standardized as above, colors are the same). What’s remarkable is that the width of the human jaw is pretty much the same from infancy to adulthood. I remember thinking this when I first started looking at human jaws early last summer, but I’d never looked at how they compare with A. robustus, whose jaw continues to increase in absolute and relative width with age (and possibly even through adulthood; Lockwood et al. 2007). This plot is admittedly a bit confusing, as sizes are measured relative to the smallest and not youngest individuals, and the narrowest human jaw is in dental stage 4. The A. robustus sample also includes a very old adult (the highest point on the plot) while the human sample only goes to early adulthood. But the basic patterns are still pretty different: A. robustus jaws get wider up to dental stage 5 (you could think of it as pre- or early adolescence) then level out (not including our large older adult), but humans’ average jaw width is fairly constant throughout ontogeny. Of course, this is at only one position along the jaw, and others will probably different.
The fragmented jaws of the youngest A. robustus (i.e. SK 63 and SK 438) do not look too different from their human counterparts, but adults are very different. Here we can see part of the reason why. Bear in mind, though, that other aspects of mandible shape do differ between these species from birth. For example, humans have a bony chin from infancy, whereas A. robustus always lacks a true chin (SK 74 is an older, probably female adult A. robustus that does have a rather anomalous “chin” but it is not homologous to ours). Not all aspects of species-specific mandible shape arise during postnatal growth!

But there you go, an enlarged human sample produces a result consistent with my earlier observation. Note that these pictures do not represent statistical tests of my hypothesis! Yes, a visual inspection of the plotted numbers suggests the two species differ in how jaw height and width grows. But saying something statistical and “definitive” is difficult. In terms of height, growth does seem pretty much the same during childhood, but then divergent later on. Width growth in the two species seems totally different. To further complicate things, a “shape” ratio of jaw width divided by height (not shown) suggests parallel (but not identical) growth trajectories in the two species. What do these observed differences mean for the null hypothesis? Which and how many variables can differ before I can feel confident about whether to reject the hypothesis? Oy, I have my work cut out for me. Stay tuned!
That paper I referenced
Lockwood, C., Menter, C., Moggi-Cecchi, J., & Keyser, A. (2007). Extended Male Growth in a Fossil Hominin Species Science, 318 (5855), 1443-1446 DOI: 10.1126/science.1149211
 Some researchers abhor cross-sectional data, pointing out that the intricacies of individuals’ longitudinal growth will not be adequately captured in with cross-sectionally. American anthropology founder Franz Boas himself discussed this in a paper nearly 82 years ago. Anyway, I was reminded of this dichotomy today when perusing a paper that examined longitudinal brain activity in a cohort of adolescent kids (right, from Campbell et al. in press). The mess of jagged lines are individuals’ measurements from age 9-18, and the smoothed blue and red curves are the cross-sectionalized curves calculated from these kids. Oy, look at all that variation and caprice that gets left out in the cross-sectionalized curves!
Some researchers abhor cross-sectional data, pointing out that the intricacies of individuals’ longitudinal growth will not be adequately captured in with cross-sectionally. American anthropology founder Franz Boas himself discussed this in a paper nearly 82 years ago. Anyway, I was reminded of this dichotomy today when perusing a paper that examined longitudinal brain activity in a cohort of adolescent kids (right, from Campbell et al. in press). The mess of jagged lines are individuals’ measurements from age 9-18, and the smoothed blue and red curves are the cross-sectionalized curves calculated from these kids. Oy, look at all that variation and caprice that gets left out in the cross-sectionalized curves!






{kind=link}