Gibbons are sometimes referred to as “lesser apes” since they’re the smaller-bodied cousins of “great apes” like us humans, chimpanzees, gorillas, and orangutans. But what they lack in body mass they make up for in taxonomic diversity, with roughly 20 species distributed across four genus groups (Kim et al., 2011). And while male great apes (except humans) have large canine teeth, both sexes in gibbons have large maxillary canines — flashy weaponry for defending territory.
Pointy canine teeth peeking out from the upper and lower jaws of an adult female gibbon cared for at the International Primate Protection League (source)
My research has generally focused on brains and growth throughout human evolution, but I started looking at gibbons a few years ago when the COVID-19 pandemic put research travel on hold. Inspired by Julia Zichello’s 2018 article about gibbon models for understanding hominin evolution and appreciating that “overlooked small apes need more attention,” I had the opportunity to CT scan a unique skeletal collection of white-handed gibbons (Hylobates lar), which was sadly harvested from the forests of Thailand back in the late 1930s. Previous research on skull growth in gibbons has mostly used small samples compiled from different species (and sometimes even different genera). In contrast, this CT dataset includes many individuals at each stage of maturation from late infancy through adulthood, effectively representing a single population at a point in time. So with this larger cross-sectional sample of a single species, we can better understand how gibbon brains and faces grow. And because permanent teeth form in a long, continuous sequence throughout the growth period, an individual’s state of dental development can serve as a marker of where they are along the maturation process.
In a paper hot off the press, Julia Boughner and I analyzed dental development in this unique sample (article here). One of the coolest things we found was that gibbons’ large upper canine teeth are among the first to begin but last to finish tooth formation. In fact, the large canines growing inside relatively small faces may inhibit growth of one of the neighboring incisor teeth until the face has grown to create enough space for it. And while most teeth developing within the jaw begin emerging into the mouth once there’s enough room for them, gibbons’ gargantuan upper canines are forced out of hiding as they outgrow their bony crypts (check out the right-most jaw in the second row below).
Cross-sectional representation of tooth formation in white-handed gibbons, starting with the youngest in the top left and ending with the oldest in the bottom right. The first permanent tooth to form and emerge, M1, is highlighted along with the canine “C.”
In addition to characterizing ‘normal’ dental development, we also observed several developmental anomalies and pathologies in the sample. Our observations corroborate previous research showing that tooth formation generally proceeds ‘as scheduled’ despite various other disturbances to development.
It remains to be seen whether early development of the canine at the cost of delayed incisor formation is a pattern unique among all the apes, since most other studies of ape tooth formation have examined the lower jaw while our study focused on the upper jaws. But the canine-incisor tradeoff that we identified sets the stage for subsequent study of skull growth in this sample, as it highlights the many factors and functions that must be coordinated during growth.
While we have several projects planned with this unique dataset, we have also published the tooth formation data that we analyzed, and the original micro-CT scans themselves will be published to the online repository Morphosource.org soon, once a few more projects are finished.
Holy crap 2015 was a big year for fossils. And how fortuitous that 2016 begins on a Fossil Friday – let’s recap some of last year’s major discoveries.
Homo naledi
Some Homo naledi mandibles in order from least to most worn teeth.
The Homo naledi sample is a paleoanthropologist’s dream – a new member of the genus Homo with a unique combination of traits, countless remains belonging to at leasta dozen individuals from infant to old adult, representation of pretty much the entire skeleton, and a remarkable geological context indicative of intentional disposal of the dead (but certainly not homicide, grumble grumble grumble…). The end of 2015 saw the announcement and uproar (often quite sexist) over this amazing sample. You can expect to see more, positive things about this amazing animal in 2016.
We’ll be presenting a bunch about Homo naledi at this year’s AAPA meeting in Hotlanta. I for one will be discussing dental development at Dinaledi- here’s a teaser:
As long as we’re talking about the AAPA meetings, my colleague David Pappano and I are organizing a workshop, “Using the R Programming Language for Biological Anthropology.” Details to come!
Lemur graveyard
Homo naledi wasn’t the only miraculously copious primate sample announced in 2015. Early last year scientists also reported the discovery of an “Enormous underwater fossil graveyard,” containing fairly complete remains of probably hundreds of extinct lemurs and other animals. As with Homo naledi, such a large sample will reveal lots of critical information about the biology of these extinct species.
Australopithecus deyiremeda
Extended Figure 1h from Haile-Selassie et al. (2015), compared with Demirjian developmental stages 6-8 . While the M1 roots look like stage 8 (complete), M2 looks like stage 7 (incomplete).
We also got a new species of australopithecus last year. Australopithecus deyiremeda had fat mandibles, a relatively short face (possibly…), and smaller teeth than in contemporaneous A. afarensis. One tantalizing thing about this discovery is that we may finally be able to put a face to the mysterious foot from Burtele, since these fossils come from nearby sites of about the same geological age. Also intriguing is the possible evidence, based on published CT images (above), that A. deyiremeda had relatively advanced canine and delayed molar development, a pattern generally attributed to Homo and not other australopithecines (if this turns out to be the case, you heard it here first!).
Lomekwian stone tool industry
3D scan and geographical location of Lomekwian tools. From africanfossils.org.
Roughly contemporaneous with A. deyiremeda, Harmand et al. (2015) report the earliest known stone tools from the 3.3 million year old site of Lomekwi 3 in Kenya. These tools are a bit cruder and much older than the erstwhile oldest tools, the Oldowan from 2.6 million years ago. These Lomekwian tools, and possible evidence for animal butchery at the 3.4 million year old Dikika site in Ethiopia (McPherron et al. 2010; Thompson et al. 2015), point to an earlier origin of lithic technology. Fossils attributed to Kenyanthropus platyops are also found at other sites at Lomekwi. With hints at hominin diversity but no direct associations between fossils and tools at this time, a lingering question is who exactly was making and using the first stone tools.
Earliest Homo
The reconstructed Ledi Geraru mandible (top left), compared with Homo naledi (top right), A. deyiremeda (bottom left), and the Uraha early Homo mandible from Malawi (bottom right). Jaws are scaled to roughly the same length from the front to back teeth; the Uraha mandible does not have an erupted third molar whereas the others do and are fully adult.
Just as Sonia Harmand and colleagues pushed back the origins of technology, Brian Villmoare et al. pushed back the origins of the genus Homo, with a 2.7 million year old mandible from Ledi Geraru in Ethiopia. This fossil is only a few hundred thousand years younger than Australopithecus afarensis fossils from the nearby site of Hadar. But the overall anatomy of the Ledi Geraru jaw is quite distinct from A. afarensis, and is much more similar to later Homo fossils (see image above). Hopefully 2016 will reveal other parts of the skeleton of whatever species this jaw belongs to, which will be critical in helping explain how and why our ancestors diverged from the australopithecines. (note that we don’t yet have a date for Homo naledi – maybe these will turn out to be older?)
Early and later Homo
Left: modified figures 2-3 from Maddux et al. (2015). Right: modified figures 7 & 13 from Ward et al. (2015). Note that in the right plot, ER 5881 femur head diameter is smaller than all other Homo except BSN 49/P27.
The earlier hominin fossil record wasn’t the only part to be shaken up. A small molar (KNM-ER 51261) and a set of associated hip bones (KNM-ER 5881) extended the lower range of size variation in Middle and Early (respectively) Pleistocene Homo. It remains to be seen whether this is due to intraspecific variation, for example sex differences, or taxonomic diversity; my money would be on the former.
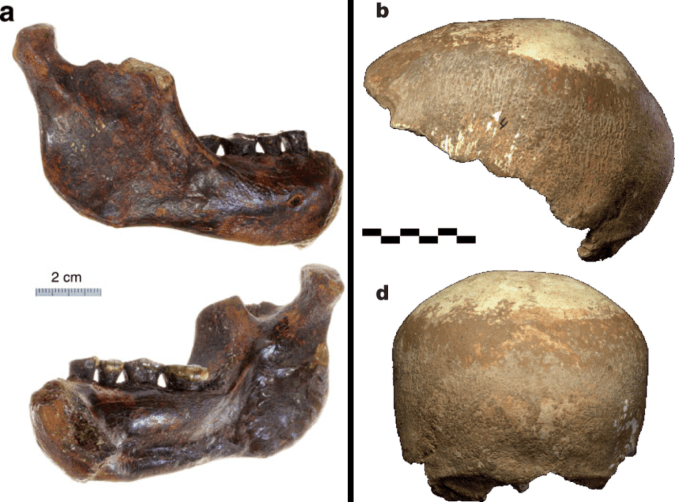
Left: Penghu 1 hemi-mandible (Chang et al. 2015: Fig. 3), viewed from the outside (top) and inside (bottom). Right: Manot 1 partial cranium (Hershkovitz et al. 2015: Fig. 2), viewed from the left (top) and back (bottom).
At the later end of the fossil human spectrum, researchers also announced an archaic looking mandible dredged up from the Taiwan Straits, and a more modern-looking brain case from Israel. The Penghu 1 mandible is likely under 200,000 years old, and suggests a late survival of archaic-looking humans in East Asia. Maybe this is a fossil Denisovan, who knows? What other human fossils are waiting to be discovered from murky depths?
The Manot 1 calvaria looks very similar to Upper Paleolithic European remains, but is about 20,000 years older. At the ESHE meetings, Israel Hershkovitz actually said the brain case compares well with the Shanidar Neandertals. So wait, is it modern or archaic? As is usually the case, with more fossils come more questions.
Crazy dinosaurs
Yi qi was bringing Skeksi back, and its upper limb had a wing-like shape not seen in any other dinosaur, bird or pterosaur. There were a number of other interesting non-human fossil announcements in 2015 (see here and here), proving yet again that evolution is far more creative than your favorite monster movie makers.
What a year – new species, new tool industries, new ranges of variation! 2015 was a great year to be a paleoanthropologist, and I’ll bet 2016 has just as much excitement in store.
References (in order of appearance)
Haile-Selassie, Y., Gibert, L., Melillo, S., Ryan, T., Alene, M., Deino, A., Levin, N., Scott, G., & Saylor, B. (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity Nature, 521 (7553), 483-488 DOI: 10.1038/nature14448
Harmand, S., Lewis, J., Feibel, C., Lepre, C., Prat, S., Lenoble, A., Boës, X., Quinn, R., Brenet, M., Arroyo, A., Taylor, N., Clément, S., Daver, G., Brugal, J., Leakey, L., Mortlock, R., Wright, J., Lokorodi, S., Kirwa, C., Kent, D., & Roche, H. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521 (7552), 310-315. DOI: 10.1038/nature14464
McPherron, S., Alemseged, Z., Marean, C., Wynn, J., Reed, D., Geraads, D., Bobe, R., & Béarat, H. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature, 466 (7308), 857-860. DOI: 10.1038/nature09248
Thompson, J., McPherron, S., Bobe, R., Reed, D., Barr, W., Wynn, J., Marean, C., Geraads, D., & Alemseged, Z. (2015). Taphonomy of fossils from the hominin-bearing deposits at Dikika, Ethiopia Journal of Human Evolution, 86, 112-135 DOI: 10.1016/j.jhevol.2015.06.013
Villmoare, B., Kimbel, W., Seyoum, C., Campisano, C., DiMaggio, E., Rowan, J., Braun, D., Arrowsmith, J., & Reed, K. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia Science, 347 (6228), 1352-1355 DOI: 10.1126/science.aaa1343
Maddux, S., Ward, C., Brown, F., Plavcan, J., & Manthi, F. (2015). A 750,000 year old hominin molar from the site of Nadung’a, West Turkana, Kenya Journal of Human Evolution, 80, 179-183 DOI: 10.1016/j.jhevol.2014.11.004
Ward, C., Feibel, C., Hammond, A., Leakey, L., Moffett, E., Plavcan, J., Skinner, M., Spoor, F., & Leakey, M. (2015). Associated ilium and femur from Koobi Fora, Kenya, and postcranial diversity in early Homo Journal of Human Evolution, 81, 48-67 DOI: 10.1016/j.jhevol.2015.01.005
Chang, C., Kaifu, Y., Takai, M., Kono, R., Grün, R., Matsu’ura, S., Kinsley, L., & Lin, L. (2015). The first archaic Homo from Taiwan Nature Communications, 6 DOI: 10.1038/ncomms7037
Hershkovitz, I., Marder, O., Ayalon, A., Bar-Matthews, M., Yasur, G., Boaretto, E., Caracuta, V., Alex, B., Frumkin, A., Goder-Goldberger, M., Gunz, P., Holloway, R., Latimer, B., Lavi, R., Matthews, A., Slon, V., Mayer, D., Berna, F., Bar-Oz, G., Yeshurun, R., May, H., Hans, M., Weber, G., & Barzilai, O. (2015). Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans Nature, 520 (7546), 216-219 DOI: 10.1038/nature14134
It will be lots of work to prep my Human Evolution course for the Fall. This past year has seen many major fossil discoveries, and adding to the list is the newly described species Australopithecus deyiremeda (Haile-Selassie et al., 2015). The fossils come from newly announced sites in Ethiopia (here it is on a map!), dating to around 3.4 million years ago. These new fossils are contemporaneous with Australopithecus afarensis, fossils attributed to Kenyanthropus platyops, and whatever the hell the Burtele foot belongs to.
The main specimens are a fairly complete half of a maxilla (upper jaw) and two decent mandibles (lower jaw bones). These fossils do not belong to the same individual (despite all the media pictures of the upper and lower jaws together). One of the most distinctive features of these fossils is how thick, both in absolute and relative terms, the mandibles are, especially given how short they are. What sticks out to me though, is that the upper jaw looks like it might have still had some growing to do. Why on earth would I think so? (The following is based off pictures from the publications, so I could be wrong!)
Extended Figure 1a from the paper, the type specimnen BRT-VP-3/1 maxilla viewed from the left side. I’ve added the M2 label for your reading pleasure.
The holotype maxilla (BRT-VP-3/1) is described as coming from a “young adult” in the Supplementary Information. However, it looks like the second molar tooth (M2) is not quite fully erupted and in occlusion, although this could be due to the natural arc of the tooth row. There is no visible wear on the tooth in the pictures, and indeed the Supplementary Information says the tooth is unworn. This means that the tooth is only recently emerged, and may not have passed the gum line, and therefore hasn’t seen much/any use yet. Authors note in the Supplementary Information that there is no M3 (a.k.a. “wisdom tooth”) wear facet on the back of M2 , meaning the last tooth hadn’t yet emerged yet either. So, this all points to a non-adult age by tooth eruption standards.
Extended Figure 1d from the paper. Same fossil, but from the bottom; pretend you’re a dentist peering into its mouth. Back is to the bottom.
In addition, the M2 roots don’t look fully formed. This is especially apparent in Extended Figure 1h, a CT section through the teeth:
Left side: Extended Figure 1h from the paper. From left to right, the teeth are P3, P4, M1, and M2. For comparison, to the right are Demirjian tooth development stages, modified from Table 2 of Kuykendall, 1996. Also compare the M2 roots with completed roots of the M1.
In many human populations, this stage of M2 development is reached (on average) between 11-13 years (Liversidge et al., 2006). In the wild Taï Forest chimpanzee sample, two individuals with M2 root completely formed (Stage H) are 10 and 11 years old (Smith et al., 2010). These apes would not be fully mature and their facial dimensions would likely have increased had they reached adulthood (Zihlman et al., 2007).
So what this suggests to me is that this maxilla may not accurately represent adult anatomy in this newly described species. In humans, the face continues to grow downwards from adolescence into adulthood, and in apes the face continues growing both forward and downward. In the differential diagnosis of A. deyiremeda, Haile-Selassie and team state, in layman’s terms, that the cheeks are positioned more toward the front than in A. afarensis, and that the front of the face doesn’t stick out as much as in A. garhi. If this specimen was not fully grown, it is likely that the true adult anatomy would have had a face that sticks out more and has less forward-positioned cheeks than in this specimen.
But, this is all speculative, and I’d like to reiterate that these observations of dental development are based only on the published pictures. Just a thought!
Two anthropology papers came out yesterday in advance print at the Proceedings of the National Academy of Sciences. I’d like first to draw your attention to the fact that they’re open access – normally such scientific papers are behind a paywall, but these two can be obtained by anyone (well, anyone with internet). One is about the chronology and nature of Acheulean technology at the 1.7-1.0 mya site of Konso in Ethiopia. The other, and the subject of this post, is about life history in wild chimpanzees from Uganda.
Tanya Smith and colleagues analyzed behavior of chimps and photographs of chimps’ erupting first molars (“M1”) to determine a] the age at which these events happen in the wild (in this population at least), and b] whether M1 eruption is tightly linked with other important life history variables, such as the adoption of adult foods, as had previously been claimed. What an adorable study – check out figure 1 from the paper (right):
Figuring out age at M1 eruption in wild, healthy chimps is important because there has been debate about whether wild chimps actually erupt their teeth at as young of ages as they do in captivity – not natural conditions. This question has recently been investigated in a skeletal sample of wild chimps of known age, from Tai forest in Cote d’Ivoire (Zihlman et al. 2004, T Smith et al. 2010), but somehow these studies raised more questions than they answered (e.g. BH Smith and Boesch 2011). So TM Smith and colleagues decided to further address this question with photographic evidence of living, arguably healthy chimps.
They found that M1 eruption occurred anywhere from 2.8-3.3 years of age in their sample of 5 cuddly infants, consistent with estimates from captivity. I have to say I’m a bit surprised it wasn’t later (but what fun is science if it’s not surprising?). Of course, this is based on 5 infants from one population, so it could stand to be reinvestigated in other chimp populations as the authors note.
Smith et al’s second task was to see how well age at M1 eruption coincided with other life history variables – this is supposed to be an important event, alleged to coincide with cessation of weaning and the adoption of adult foods. Moreover, since a mother is no longer nursing her infant, M1 eruption “should” also be roughly contemporaneous with a mother’s return to estrus cycling and subsequent reproduction. Many infants were observed to begin eating adult-like foods prior to M1 eruption, around 3 years. Unexpectedly however, infants also nursed for a while even after M1 eruption. In fact, time spent nursing actually increased for a brief period around 3 years of age, possibly because their mothers’ milk was not as nutritious as at younger ages.
Now, what interests me most about this are possible implications for the evolution of growth and life history. Many researchers have argued that extinct hominids, like the australopithecines, would have grown up relatively rapidly like apes, rather than slowly like humans. This claim has been based pretty much entirely on dental development, until my dissertation research. There, I’ve shown that one hominid, Australopithecus robustus, probably experienced greater jaw growth than humans prior to eruption of the M2. Now, if this hominid erupted its teeth as fast as apes, and grew more than humans, this implies really really high growth rates for A. robustus (that is, if we can extrapolate from the jaw to the overall body size).
I’ll be working a bit more on this latter point in the near future. In the mean time, let’s hear it for open access bioanthro Continue reading →