Homo habilis just got some long arms to go along with its dexterous hands. In a recent paper in the journal The Anatomical Record, Fred Grine and colleagues describe and analyze some spectacular fossils recovered near the town of Ileret in Kenya, dating to just over 2 million years ago. There were a few different kinds of human-like species inhabiting the planet around this time, but researchers were able to assign these bones to Homo habilis thanks to some chemical clues connecting them to a nearly complete set of teeth found a few meters away. This partial skeleton of a young adult individual is an incredible discovery, connected by clever scientific sleuthing, and provides important information about an early member of the human lineage.
You can see some great photos of these fossils (as well as a fantastic fossil foot of a different individual) in a 2015 press release from the Turkana Basin Institute. A more recent announcement from the Institut Català Paleontologia includes a photo showing the late great Bill Jungers and fossil maven Meave Leakey with the fossils, which helps show the actual size of the bones.
Ann Gibbons’ article about the discovery has a great quote from paleoanthropologist Stephanie Melillo (who discovered the Burtele foot fossil): “If you dressed up a Homo habilis individual in clothes and you saw her walking in the distance, would you do a double take? This study shows us that the answer is YES!”
Artist’s depiction of Homo habilis dressed up in clothes and you see her walking in the distance (image source)
The reason we might react to seeing Homo habilis like Gertie glimpsing E.T., as this skeleton shows, is that this early human had longer arms (especially forearms) than most of us do today. Thickness of the bones also shows that they were probably quite strong as a result of experiencing lots of force from use during life. Long and strong hominin arms are typically interpreted as evidence that these ancient ancestors spent a good deal of time climbing trees.
These features have previously been documented in some of the few other partial skeletons attributed to Homo habilis, as Grine and colleagues note. Indeed, the new article does a deep dive into what is known (and unknown) about the bones and body of Homo habilis, and it also provides a thoughtful review of recent research cautioning against over-interpreting climbing behaviors from fossil remains.
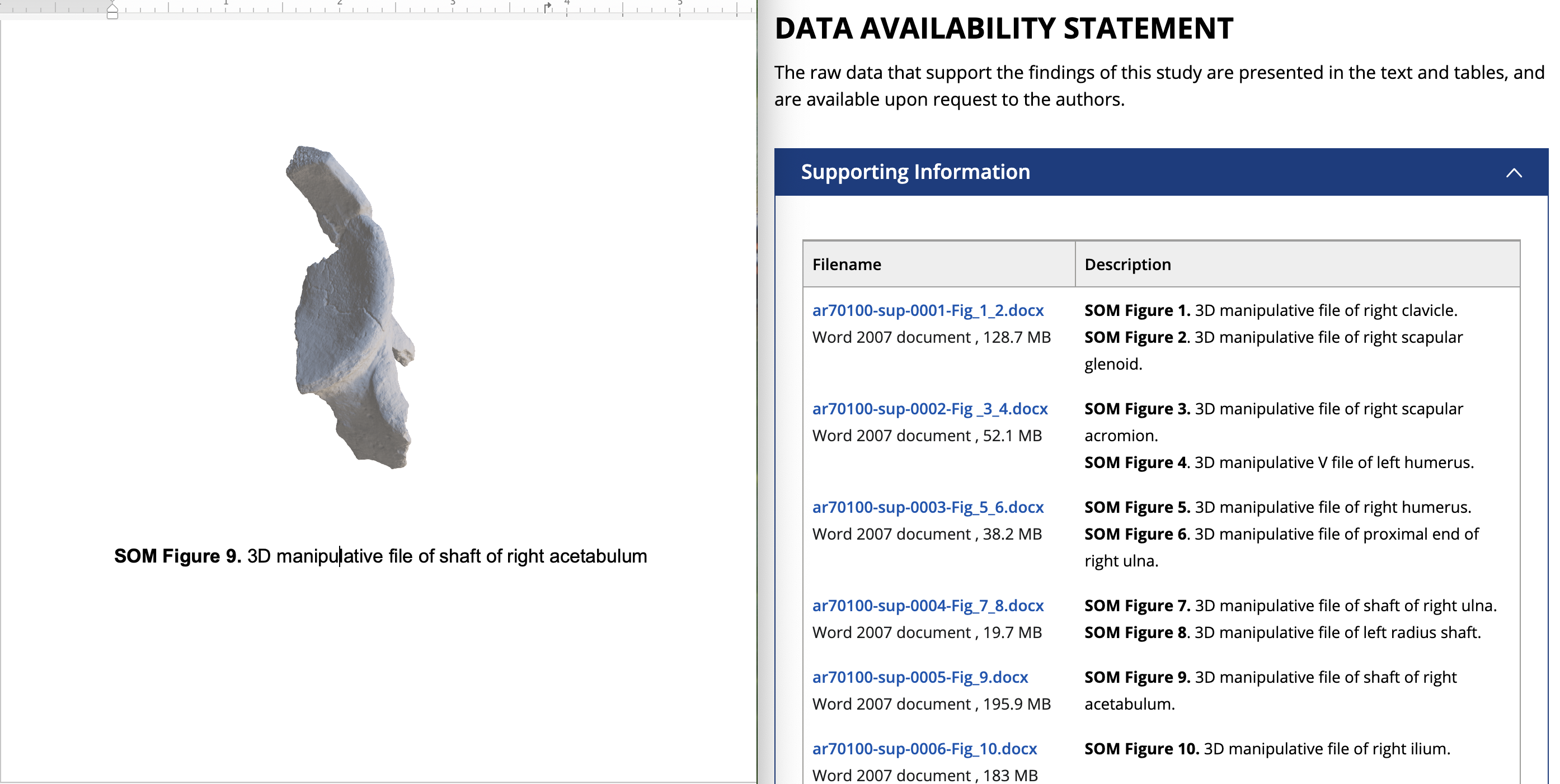
For more fossil fun, the article’s supporting online material includes “3D manipulative files” of the original specimens, so anyone can have a look at the bones in 3D using Microsoft Word:
Dr. Yohannes Haile-Selassie & colleagues just published some amazing fossils from around 3.4 million years ago, that convincingly link an unusual hominin foot fossil to an ancient human called Australopithecus deyiremeda.
In 2012, Haile-Selassie and team reported a foot fossil from Burtele, Ethiopia, revealing a bipedal creature (like a human) but with some grasping ability in the big toe (like all other primates). Then in 2015, the team presented some jaws and teeth from a nearby geological locality in the Burtele region, around which they designated a new hominin species, Australopithecus deyiremeda. The researchers hesitated to allocate the Burtele foot to this new species since they didn’t have similar fossils for comparison between the different fossil localities. But as the scientists have recently reported, jaws and teeth discovered from the foot site, including an incredible juvenile mandible, match those of Au. deyiremeda from the nearby Burtele sites. Now we can put a foot to the name.
The Burtele fossils help reveal the diversity of early hominins like Australopithecus and the contexts out of which our own genus Homo evolved. What caught my attention hiding among this amazing assemblage was a fossil that only gets a quick mention in the paper—the ischium bone from the hip of a juvenile deyiremeda:
Extended Data Figure 7 from Haile-Selassie et al. (2025). The BRT-VP-2/87 juvenile ischium (from the right side of the body), depicted in side (a), middle (b), and back (c) views.
The fossil, given the catalog number BRT-VP-2/87, represents a different individual from the juvenile jaw mentioned above. It nevertheless provides a great deal of information despite being a small fragment (less than 2 inches long). The authors observe that the body of the ischium that extends beneath the hip joint is quite long, similar to modern apes, fossil Ardipithecus ramidus, and australopiths. This contrasts with the ischium of modern and fossil Homo in which the bone projects less beyond the hip socket:
Right juvenile ischium bones, scaled to similar size and oriented in similar positions. The black line on each depicts the distance from the hip socket margin to the top of the ischial tuberosity (left modified from Scheuer & Black, 2000 Fig. 10.15)
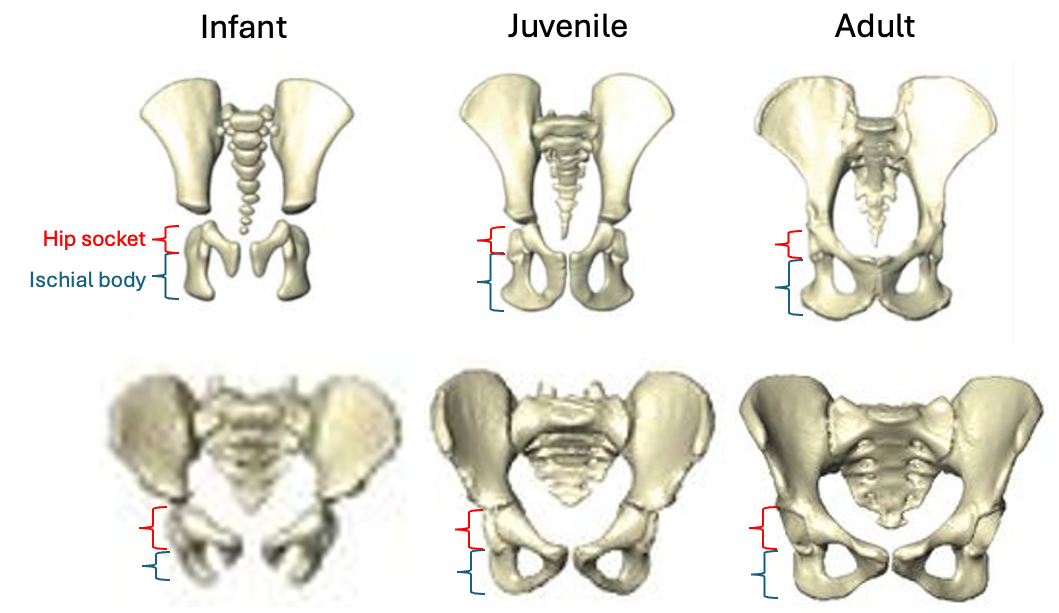
The bottom of the ischium is called the “ischial tuberosity,” and is the attachment surface for the hamstrings muscles. Having a long ischium provides the hamstrings of apes and other arboreal primates with more powerful hip extension—very useful when climbing trees but it also limits how far back the thigh can extend away from the body (Kozma et al., 2018). The shorter ischium of humans, Homo naledi, and other members of our genus may make our hamstrings a little less powerful, but it also helps us fully extend our legs which is crucial to our efficient bipedal walking and running.
Pelvis growth and development in chimpanzees (top row) and humans (bottom row), all scaled to a similar vertical height. Notice the differences in both the relative length of the ischium (blue bracket) and orientation of the ischial tuberosities between chimps and humans, consistent across the growth period. Images modified from Huseynov et al. (2016 and 2017).
Based on studies of modern humans and other primates, we know that this configuration of bones and muscles is established before birth, so we can be confident that adult Au. deyiremeda would have had a similar anatomy to BRT-VP-2/73, albeit at an unknown, larger size. A hip well adapted for climbing is consistent with the Burtele foot with a grasping big toe.
As Haile-Selassie and colleagues note in the online supplementary information accompanying the paper, only immature fossils allow us to reconstruct the evolution of growth and development. But one of the major challenges of studying immature remains is determining their age or state of maturation, which is critical for understanding how much change occurs between, say, infancy and adulthood. The authors of this study note that the qualitative appearance of the BRT-VP-2/73 hip socket surface is like that of modern humans around 6 years of age, yet the fossil is much smaller and more similar in size to 3 year-old humans. My colleagues and I (2022) faced a similar challenge when analyzing a juvenile Homo naledi hip, and we also relied on qualitative comparisons of how the joint “looks” at different stages of development.
But I think we’re at a point now where we can try to quantify some of these tricky developing surfaces to help place immature fossils more precisely along a timeline of development. For example, Peter Stamos & Tim Weaver (2020) adapted a method for quantifying the topography of teeth, to measure the complex curvature of the developing surface of the knee. If these quantitative methods can distinguish different phases of development in large samples of humans and other primates (e.g., Stamos et al., 2025), they could then be extended to the immature hominin fossil record.
Some cool insights could also be gained by applying older and established methods like landmark-based geometric morphometrics, even on quite fragmentary fossils. This approach could capture the development and orientation of the ischial tuberosity relative to the hip socket surface in fragments like BRT-VP-2/73, MLD 8, and Homo naledi fossils (depicted above) and compared with fossil adults. Researchers have also devised robust ways of quantifying size and shape changes during growth based on modern animals, and using these patterns to then ‘grow’ immature fossils to more developed states, for comparison with actual adult fossils (McNulty et al., 2006). Applying this approach to even just the small fossil sample of ischia described here could tell us a lot about how ancient animals moved at different periods in their lives. Someone just needs to park their ischial tuberosities in a chair and do it!
A growing fossil record of immature hominins, alongside technical advances in quantifying and comparing anatomy, mean that we are ready to learn much more about how our extinct ancestors and cousins grew into competent adults.
Holy crap 2015 was a big year for fossils. And how fortuitous that 2016 begins on a Fossil Friday – let’s recap some of last year’s major discoveries.
Homo naledi
Some Homo naledi mandibles in order from least to most worn teeth.
The Homo naledi sample is a paleoanthropologist’s dream – a new member of the genus Homo with a unique combination of traits, countless remains belonging to at leasta dozen individuals from infant to old adult, representation of pretty much the entire skeleton, and a remarkable geological context indicative of intentional disposal of the dead (but certainly not homicide, grumble grumble grumble…). The end of 2015 saw the announcement and uproar (often quite sexist) over this amazing sample. You can expect to see more, positive things about this amazing animal in 2016.
We’ll be presenting a bunch about Homo naledi at this year’s AAPA meeting in Hotlanta. I for one will be discussing dental development at Dinaledi- here’s a teaser:
As long as we’re talking about the AAPA meetings, my colleague David Pappano and I are organizing a workshop, “Using the R Programming Language for Biological Anthropology.” Details to come!
Lemur graveyard
Homo naledi wasn’t the only miraculously copious primate sample announced in 2015. Early last year scientists also reported the discovery of an “Enormous underwater fossil graveyard,” containing fairly complete remains of probably hundreds of extinct lemurs and other animals. As with Homo naledi, such a large sample will reveal lots of critical information about the biology of these extinct species.
Australopithecus deyiremeda
Extended Figure 1h from Haile-Selassie et al. (2015), compared with Demirjian developmental stages 6-8 . While the M1 roots look like stage 8 (complete), M2 looks like stage 7 (incomplete).
We also got a new species of australopithecus last year. Australopithecus deyiremeda had fat mandibles, a relatively short face (possibly…), and smaller teeth than in contemporaneous A. afarensis. One tantalizing thing about this discovery is that we may finally be able to put a face to the mysterious foot from Burtele, since these fossils come from nearby sites of about the same geological age. Also intriguing is the possible evidence, based on published CT images (above), that A. deyiremeda had relatively advanced canine and delayed molar development, a pattern generally attributed to Homo and not other australopithecines (if this turns out to be the case, you heard it here first!).
Lomekwian stone tool industry
3D scan and geographical location of Lomekwian tools. From africanfossils.org.
Roughly contemporaneous with A. deyiremeda, Harmand et al. (2015) report the earliest known stone tools from the 3.3 million year old site of Lomekwi 3 in Kenya. These tools are a bit cruder and much older than the erstwhile oldest tools, the Oldowan from 2.6 million years ago. These Lomekwian tools, and possible evidence for animal butchery at the 3.4 million year old Dikika site in Ethiopia (McPherron et al. 2010; Thompson et al. 2015), point to an earlier origin of lithic technology. Fossils attributed to Kenyanthropus platyops are also found at other sites at Lomekwi. With hints at hominin diversity but no direct associations between fossils and tools at this time, a lingering question is who exactly was making and using the first stone tools.
Earliest Homo
The reconstructed Ledi Geraru mandible (top left), compared with Homo naledi (top right), A. deyiremeda (bottom left), and the Uraha early Homo mandible from Malawi (bottom right). Jaws are scaled to roughly the same length from the front to back teeth; the Uraha mandible does not have an erupted third molar whereas the others do and are fully adult.
Just as Sonia Harmand and colleagues pushed back the origins of technology, Brian Villmoare et al. pushed back the origins of the genus Homo, with a 2.7 million year old mandible from Ledi Geraru in Ethiopia. This fossil is only a few hundred thousand years younger than Australopithecus afarensis fossils from the nearby site of Hadar. But the overall anatomy of the Ledi Geraru jaw is quite distinct from A. afarensis, and is much more similar to later Homo fossils (see image above). Hopefully 2016 will reveal other parts of the skeleton of whatever species this jaw belongs to, which will be critical in helping explain how and why our ancestors diverged from the australopithecines. (note that we don’t yet have a date for Homo naledi – maybe these will turn out to be older?)
Early and later Homo
Left: modified figures 2-3 from Maddux et al. (2015). Right: modified figures 7 & 13 from Ward et al. (2015). Note that in the right plot, ER 5881 femur head diameter is smaller than all other Homo except BSN 49/P27.
The earlier hominin fossil record wasn’t the only part to be shaken up. A small molar (KNM-ER 51261) and a set of associated hip bones (KNM-ER 5881) extended the lower range of size variation in Middle and Early (respectively) Pleistocene Homo. It remains to be seen whether this is due to intraspecific variation, for example sex differences, or taxonomic diversity; my money would be on the former.
Left: Penghu 1 hemi-mandible (Chang et al. 2015: Fig. 3), viewed from the outside (top) and inside (bottom). Right: Manot 1 partial cranium (Hershkovitz et al. 2015: Fig. 2), viewed from the left (top) and back (bottom).
At the later end of the fossil human spectrum, researchers also announced an archaic looking mandible dredged up from the Taiwan Straits, and a more modern-looking brain case from Israel. The Penghu 1 mandible is likely under 200,000 years old, and suggests a late survival of archaic-looking humans in East Asia. Maybe this is a fossil Denisovan, who knows? What other human fossils are waiting to be discovered from murky depths?
The Manot 1 calvaria looks very similar to Upper Paleolithic European remains, but is about 20,000 years older. At the ESHE meetings, Israel Hershkovitz actually said the brain case compares well with the Shanidar Neandertals. So wait, is it modern or archaic? As is usually the case, with more fossils come more questions.
Crazy dinosaurs
Yi qi was bringing Skeksi back, and its upper limb had a wing-like shape not seen in any other dinosaur, bird or pterosaur. There were a number of other interesting non-human fossil announcements in 2015 (see here and here), proving yet again that evolution is far more creative than your favorite monster movie makers.
What a year – new species, new tool industries, new ranges of variation! 2015 was a great year to be a paleoanthropologist, and I’ll bet 2016 has just as much excitement in store.
References (in order of appearance)
Haile-Selassie, Y., Gibert, L., Melillo, S., Ryan, T., Alene, M., Deino, A., Levin, N., Scott, G., & Saylor, B. (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity Nature, 521 (7553), 483-488 DOI: 10.1038/nature14448
Harmand, S., Lewis, J., Feibel, C., Lepre, C., Prat, S., Lenoble, A., Boës, X., Quinn, R., Brenet, M., Arroyo, A., Taylor, N., Clément, S., Daver, G., Brugal, J., Leakey, L., Mortlock, R., Wright, J., Lokorodi, S., Kirwa, C., Kent, D., & Roche, H. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521 (7552), 310-315. DOI: 10.1038/nature14464
McPherron, S., Alemseged, Z., Marean, C., Wynn, J., Reed, D., Geraads, D., Bobe, R., & Béarat, H. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature, 466 (7308), 857-860. DOI: 10.1038/nature09248
Thompson, J., McPherron, S., Bobe, R., Reed, D., Barr, W., Wynn, J., Marean, C., Geraads, D., & Alemseged, Z. (2015). Taphonomy of fossils from the hominin-bearing deposits at Dikika, Ethiopia Journal of Human Evolution, 86, 112-135 DOI: 10.1016/j.jhevol.2015.06.013
Villmoare, B., Kimbel, W., Seyoum, C., Campisano, C., DiMaggio, E., Rowan, J., Braun, D., Arrowsmith, J., & Reed, K. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia Science, 347 (6228), 1352-1355 DOI: 10.1126/science.aaa1343
Maddux, S., Ward, C., Brown, F., Plavcan, J., & Manthi, F. (2015). A 750,000 year old hominin molar from the site of Nadung’a, West Turkana, Kenya Journal of Human Evolution, 80, 179-183 DOI: 10.1016/j.jhevol.2014.11.004
Ward, C., Feibel, C., Hammond, A., Leakey, L., Moffett, E., Plavcan, J., Skinner, M., Spoor, F., & Leakey, M. (2015). Associated ilium and femur from Koobi Fora, Kenya, and postcranial diversity in early Homo Journal of Human Evolution, 81, 48-67 DOI: 10.1016/j.jhevol.2015.01.005
Chang, C., Kaifu, Y., Takai, M., Kono, R., Grün, R., Matsu’ura, S., Kinsley, L., & Lin, L. (2015). The first archaic Homo from Taiwan Nature Communications, 6 DOI: 10.1038/ncomms7037
Hershkovitz, I., Marder, O., Ayalon, A., Bar-Matthews, M., Yasur, G., Boaretto, E., Caracuta, V., Alex, B., Frumkin, A., Goder-Goldberger, M., Gunz, P., Holloway, R., Latimer, B., Lavi, R., Matthews, A., Slon, V., Mayer, D., Berna, F., Bar-Oz, G., Yeshurun, R., May, H., Hans, M., Weber, G., & Barzilai, O. (2015). Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans Nature, 520 (7546), 216-219 DOI: 10.1038/nature14134
We’re learning about the divergence between robust Australopithecus and early Homo 2.5-ish million years ago in my Human Evolution class this week. Because of this multiplicity of contemporaneous species, when scientists find new hominin fossils in Early Pleistocene sites, a preliminary question is, “What species is it?”
Scrutinizing the fossil record, asking the difficult questions. (Science credit)
To help my students learn how we know whether certain fossils belong to the same species, and to which group new fossils might belong, in this week’s lab we compared tooth sizes of Australopithecus boisei and early Homo. After seeing how tooth sizes differed between these groups, students then tested whether they could determine whether two “mystery” fossils (KNM-ER 60000 and 62000; Leakey et al. 2012) belonged either group.
Early Pleistocene hominin fossils from Kenya. Left to right: KNM-ER 406, ER 62000 and ER 1470. At the center is one f the lab’s “mystery jaws.”
Students downloaded 3D scans of hominin fossils from AfricanFossils.org, and measured buccolingual/labiolingual tooth crown diameters using MeshLab.
Early Pleistocene hominin mandibles. Left to right: KNM-ER 3230, ER 60000 (“mystery” jaw) and ER 1802.
The first purpose of this lab was to help familiarize students with skull and tooth anatomy of early Pleistocene humans. Although lectures and readings are full of images, a lab activity forces students to spend time visually examining fossils. Plus, they’re in 3D which is a whole D greater than 2D – the visual equivalent of going to eleven! The second goal of the lab was to help prepare students for their term projects, in which they must pose a research question about human evolution, generate predictions, and find and use data to test hypotheses.
If you’re interested in using or adapting this activity for your class, here are the handout and data sheet into which students enter their measurements. The data sheet specifies the fossils that can be downloaded from africanfossils.org. Some relevant fossils (i.e., KNM WT 15000 and ER 992) were not included because the 3D scans yield larger measurements than in reality.
Reference Leakey MG, Spoor F, Dean MC, Feibel CS, Antón SC, Kiarie C, & Leakey LN (2012). New fossils from Koobi Fora in northern Kenya confirm taxonomic diversity in early Homo. Nature, 488 (7410), 201-4 PMID: 22874966
Figure 3 from Ward et al. 2015. A little distal to the hip, yes, but the pun still works. Views are, going clockwise starting at the top the top left, from above, from below, from the back, from the side, and from the front.
There’s also a partial ilium associated with the femur – that makes a pretty complete hip!
Figure 5 from Ward et al. shows the fossil. Jump for joy that it’s complete enough for us to tell it comes from the left side!
Despite how fragmentary the femur and ilium are, the researchers were able to estimate the diameter of the femur head and hip socket reliably. The hip joints are smaller than all Early Pleistocene Homo except for the Gona pelvis. Comparing ER 5881 the large contemporaneous KNM-ER 3228 hip bone, the authors found these two specimens to be more different in size than is usually seen between sexes of many primate species. The size difference best matches male-female differences in highly dimorphic species like gorillas.
Ward et al. find that the specimen generally looks like early Homo but that the inferred shape of the pelvic inlet is a little different from all other Early and Middle Pleistocene human fossils. The authors take this discrepancy to suggest that there was more than one “morphotype” (‘kind of shape’), and therefore possibly species, of Homo around 1.9 million years ago. While I wouldn’t just yet go so far as to say this anatomy is due to species differences, I do agree that KNM ER 5881 helps our understanding and appreciation of anatomical variation in our early ancestors. Like all great fossil discoveries, the more we find, the more we learn that we don’t know. Here’s to more Homo hips in the near future!
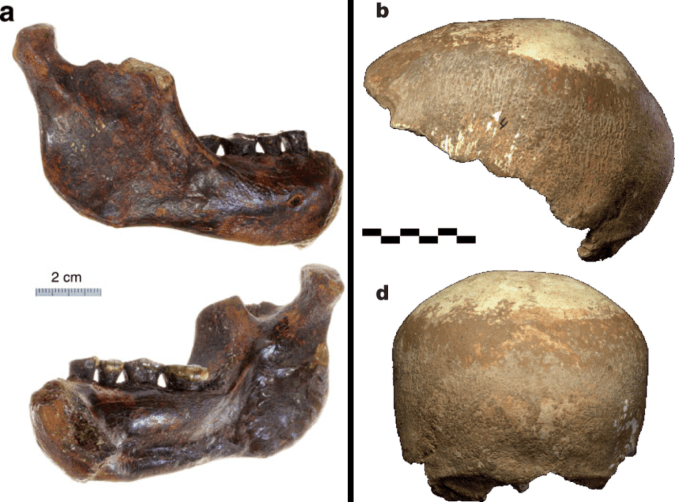
This was a big week for Middle-Late Pleistocene fossil humans. Chun-Hsiang Chang and colleagues describe a mandible dredged up off the western coast of Taiwan, which they note in the title as, “The first archaic Homo” fossil known from the region. The geological context makes it difficult to date the specimen precisely, but authors argue it is probably younger than 190 thousand years old.
The Penghu mandible. Figure 3. From Chang et al.
In life, this individual was fully grown but appears never to have developed third molars (the “wisdom teeth”). Such “third molar agenesis” is relatively rare before modern times, but is also seen in the D2735Homo erectus mandible from Dmanisi. I wouldn’t make much of this coincidence, but it does raise the question of whether the cause of agenesis, not uncommon today, was the same then as now.
Shortly after the announcement of the Penghu mandible, Israel Hershkovitz and colleagues presented a 55,000 year old brain case from Manot Cave in the Levant. The calvaria (fancy word for brain case) looks very similar to the skulls of the slightly younger “anatomically modern” humans of the Upper Paleolithic in Europe, albeit with a few Neandertal-like traits here and there (hey, just like many of the Upper Paleolithic humans).
The Manot calvaria (Figure 2 from Hershkovitz et al.) The views are (a-d) from the top with front to the left; from the left; from the front; and from the back. Extra credit: In the top view (a), can you identify the features telling that the front is to the left?
John Hawks has good posts dedicated to both Penghu and Manot. The upshot of these discoveries is that Middle and Late Pleistocene human population diversity, and the interactions between these populations, are probably much more complicated and interesting than the old model of ‘modern’ humans arising singly in Africa and replacing ‘archaic’ humans in different parts of the globe. With the technological advances and fossil discoveries of the past decade, the rather simple Replacement model has given way to a better appreciation of true complexity of human evolution toward the end of the Ice Age. Both of these new papers reflect this new perspective.
Along these lines, accompanying the Manot paper in Nature is an editorial, “Human history defies easy stories.” What caught my attention reading this (anonymous?) commentary is that it puts scientific interpretations of the past into a social and historical context. The author notes that the traditional story of modern humans arising, spreading and eradicating other groups of human has “imperialist framing, in which evolution and replacement can be justified after the fact as a kind of manifest destiny.” Science doesn’t occur in a vacuum, it’s done by people whose minds and creativities are molded in specific historical, economic and cultural contexts. This editorial comment makes one wonder how the human fossil record would have been interpreted, had most of it not discovered against the social backdrop of ruthless capitalism.
It’s been a quiet month here at Lawnchair, as I’ve just returned from the Rising Star Workshop, taking part in the analysis and description of new hominin remains from South Africa. We’ll have some exciting announcements to make in the near future.
Also, I petted a ferocious, bloodthirsty lion!
To ease back into the Lawnchair, I thought I’d resurrect eFfing Fossil Friday, a short-lived series from when I was collecting data for my dissertation three years ago (speaking of which, a paper related to my dissertation came out in AJPA during the Workshop, as well). A lot has happened since the last installment of FFF, so whose heads will be on the chopping block today?
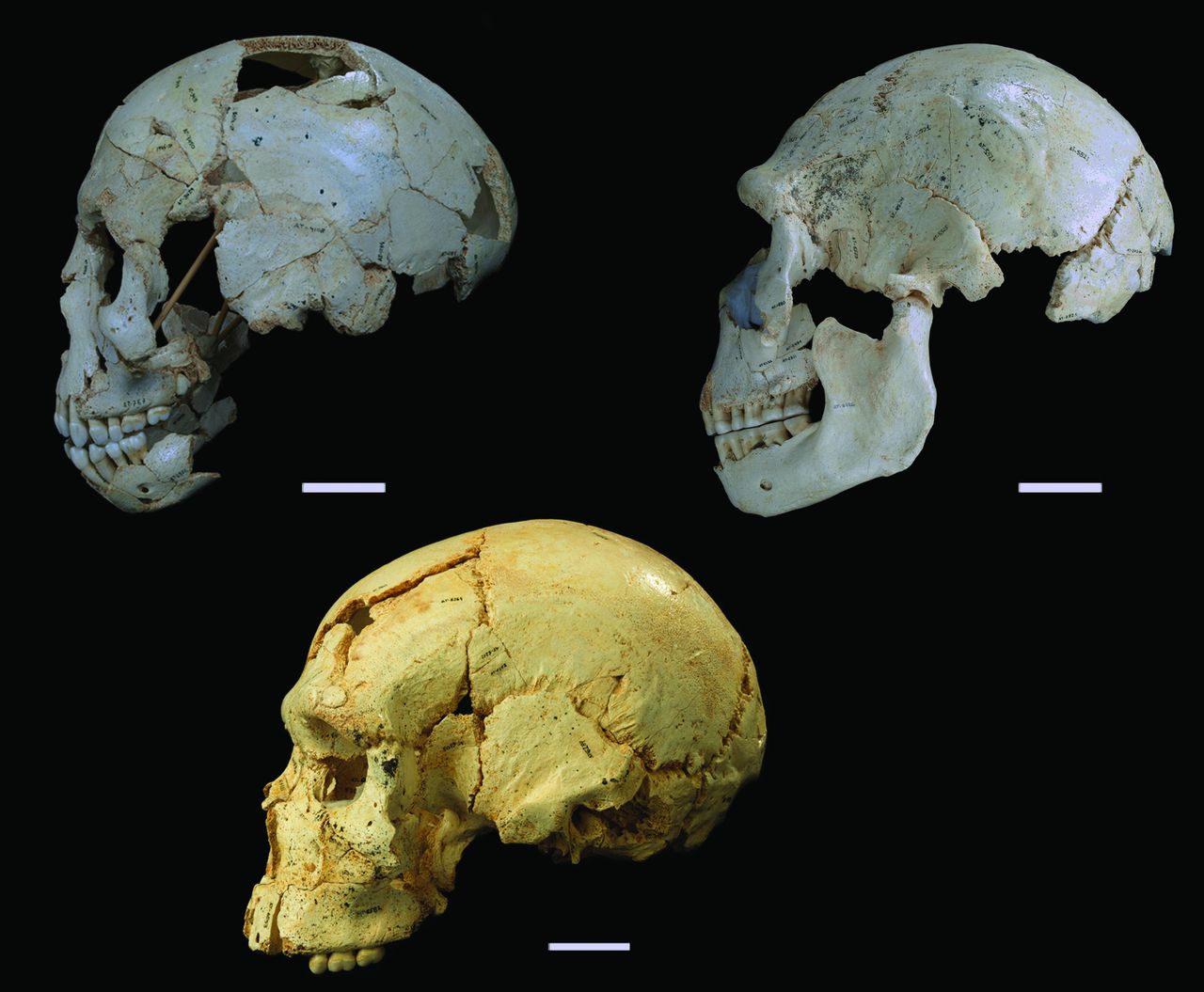
Crania #s 9, 15 and 17 (clockwise from top left). Cranium 9 is an early adolescent and the other two are adults – lookit how the facial anatomy changes with age! (Fig. 1 from Arsuaga et al., 2014)
It’s new crania from Sima de los Huesos, Atapuerca! These are published today in the journal Science by Juan L. Arsuaga and colleagues. Sima de los Huesos is a pretty remarkable site in Spain dating to the Middle Pleistocene; the site is probably at least 400,000 years old, and the remains of at least 28 individuals. These specimens show many similarities with Neandertals who later inhabited the area, but don’t have all of the ‘classic’ Neandertal features.
What I like about this figure from the paper is that the comparison of the adolescent (top left) with adults (the other two) shows how the skull changes during growth. The major visible difference is that the face sticks out in front of the brain case more in the adults than the adolescent. As a result, the adolescent lacks a supraorbital torus (“brow ridge”), but this would have developed as the face grew forward and away from the brain. Ontogeny!
It’s a new year, and while my mind should be marred by a hangover, instead all I can think about are fossils and scapulas.
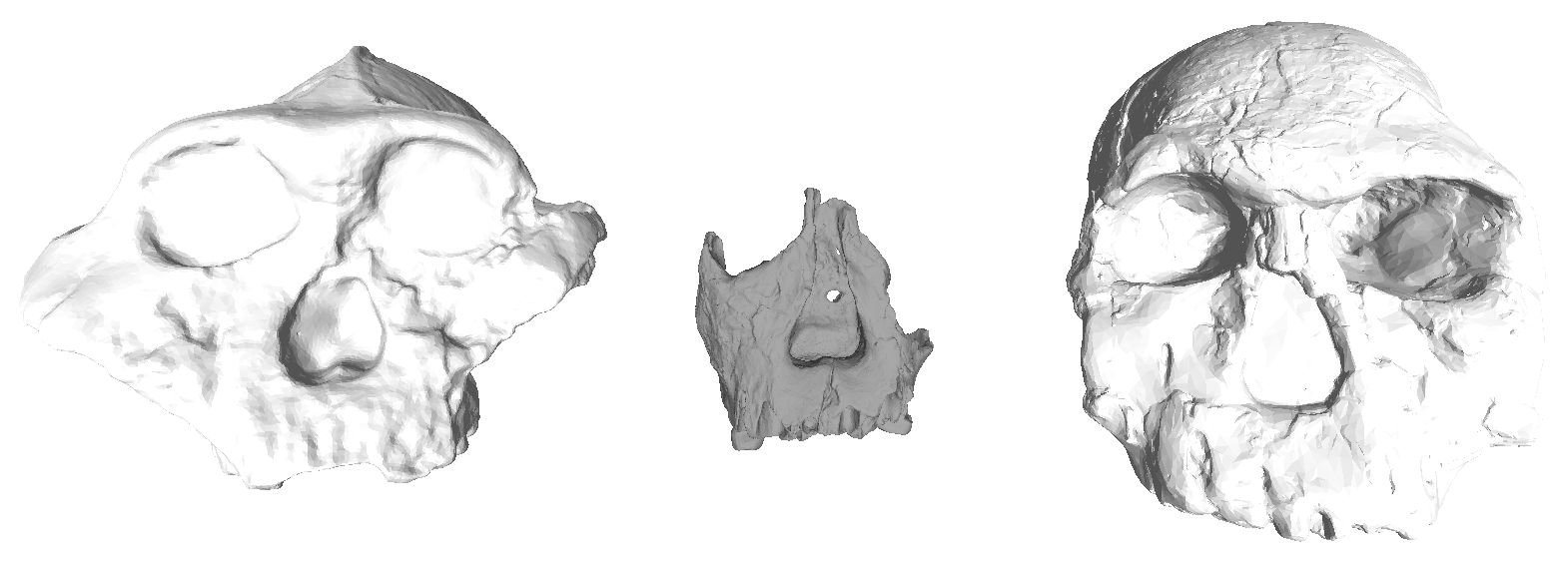
A pretty cool study was published online in the Journal of Human Evolution last week, and I’ve finally gotten to peruse it. Fabio Di Vincenzo and colleagues analyzed the shape of the outline of the glenoid fossa on the scapula (not to be confused with the glenoid on your skull), from Australopithecus africanus to present day humans. The glenoid fossa is essentially the socket in the ball-and-socket joint of your shoulder. The authors found that there is pretty much a single trend of glenoid shape change from Australopithecus through the evolution of the genus Homo: from the fairly narrow joint in Australopithecusafricanus and A. sediba, to the relatively wide joint in recent humans. The overall size and shape of the joint influences/reflects shoulder mobility, so presumably this shape change hints that more front-to-back arm motions became more important through the course of human evolution (authors suggest throwing in humans from the Late Pleistocene onward).
The finding of a single predominant trend in glenoid shape evolution is pretty interesting. On top of that, the authors add an ‘evo-devo’ twist by comparing species’ average “shapes” (first principle component scores, on the y-axis in the figure at right) with their estimated ages at skeletal maturity (which appears scaled to the modern human value, on the x-axis). Though it’s not an ideal dataset for running a linear regression, the figure at right shows that there appears to be a fairly linear relationship across human evolution, such that groups with an older age at skeletal maturity tend to have a more rounded (modern human-like) glenoid fossa (note that the individuals in the analysis were all adults). Overall size does not contribute to shape variation among these glenoids.
This work raises two issues, and ultimately leads to a testable evo-devo hypothesis. The first issue is to what extent we can trust their estimates of age at skeletal maturity. These estimates were allegedly taken from a chapter by Helmut Hemmer (2007) in the prohibitively expensive Handbook of Paleoanthropology. Cursorily glancing at this chapter, I can’t find age at skeletal maturation estimated for any hominids. It is possible that in my skimming I missed the estimates, or alternatively that Di Vincenzo and colleagues misinterpreted another variable as skeletal development. Either way, these estimates would still need to be taken with a grain of salt, given that it is almost impossible to know the true age at death of a fossil (but see Antoine et al. 2008), especially if there are no associated cranio-dental elements.
That said, it is perfectly reasonable to suppose that the age at skeletal maturation has increased over the course of human evolution; life-span increased through human evolution, and so all else being equal (which it almost certainly isn’t) we could expect that maturation would occur later over time, too. So this leads to a second issue: given the “evo-devo change” the authors hypothesize, what is the evo-devo mechanism? That is, how was development modified to effect the evolutionary changes we see in the hominid scapula? Because they found adult glenoid shape correlates with estimated age at skeletal maturity, this leads to the hypothesis that postnatal skeletal growth accounts for the shape difference. Indeed, they state:
“If functional and static allometric influences are unlikely, we…interpret the trend…as reflecting growth and developmental factors. A major, albeit gradual, trend of ontogenetic heterochrony occurred in the evolution of the genus Homo… and thus differences within and between taxa in overall growth rates may have produced the pattern of variation between samples, as well as the overall temporal trend observed. The regression of life history variables [they only looked at 1]… with PCA [principle components analysis] scores supports this ‘ontogenetic’ hypothesis.”
The authors suggest that humans’ slower growth rates but longer growth period “led to longer periods of bone deposition along the inferior-lateral edge of the [glenoid fossa]” The heterochronic process they suggest is “peramorphosis” – the descendant reaches a shape that is ‘beyond’ that of the ancestor.
The figure above is from a seminal “heterochrony” paper by Pere Alberch and colleagues (1979), portraying how peramorphosis can occur. In each, the y-axis represents shape and the x-axis is age. A the descendant’s peramorphic shape (“Ya”) could result from accelerated growth (left graph) or from an extension of growth to later ages than in the ancestor (right graph).
And so this leads to a testable hypothesis. Di Vincenzo and colleagues cite (dental) evidence that humans’ overall body growth rates are slower than earlier hominids’, undermining the hypothesis that acceleration is responsible for humans’ glenoid peramorphosis. Rather, they hypothesize that humans’ slower growth rates coupled with a longer period of skeletal development, to result in a relatively wider glenoid, due to increased development of the secondary growth centers (e.g. the graph at right, above). This developmental scenario predicts that subadult human glenoids should resemble earlier hominid adults’, that “ontogeny recapitulates phylogeny” as far as glenoid shape is concerned. Analyzing glenoid growth can even be extended to include fossils – the >3 million year old human ancestor Australopithecus afarensis has glenoids preserved for an infant (DIK-VP-1; Alemseged et al. 2006) and 2 adults (AL 288 “Lucy” and KSD-VP-1; Johanson et al. 1982, Haile-Selassie et al. 2010). An alternate hypothesis is that species’ distinct glenoid shapes are established early during life (i.e. in utero), and/or that no simple heterochronic process is involved.
Di Vincenzo’s and colleagues’ study points to the importance of development in understanding human evolution, and their hypothesized “evo-devo change” in glenoid shape is ripe for testing.
References Pere Alberch, Stephen Jay Gould, George F. Oster, & David B. Wake (1979). Size and shape in ontogeny and phylogeny Paleobiology, 5 (3), 296-317
Alemseged, Z., Spoor, F., Kimbel, W., Bobe, R., Geraads, D., Reed, D., & Wynn, J. (2006). A juvenile early hominin skeleton from Dikika, Ethiopia Nature, 443 (7109), 296-301 DOI: 10.1038/nature05047
Antoine, D., Hillson, S., & Dean, M. (2009). The developmental clock of dental enamel: a test for the periodicity of prism cross-striations in modern humans and an evaluation of the most likely sources of error in histological studies of this kind Journal of Anatomy, 214 (1), 45-55 DOI: 10.1111/j.1469-7580.2008.01010.x
Di Vincenzo, F., Churchill, S., & Manzi, G. (2011). The Vindija Neanderthal scapular glenoid fossa: Comparative shape analysis suggests evo-devo changes among Neanderthals Journal of Human Evolution DOI: 10.1016/j.jhevol.2011.11.010
Haile-Selassie, Y., Latimer, B., Alene, M., Deino, A., Gibert, L., Melillo, S., Saylor, B., Scott, G., & Lovejoy, C. (2010). An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia Proceedings of the National Academy of Sciences, 107 (27), 12121-12126 DOI: 10.1073/pnas.1004527107
Hemmer, Helmut (2007). Estimation of Basic Life History Data of Fossil Hominoids Handbook of Paleoanthropology, 587-619 DOI: 10.1007/978-3-540-33761-4_19
Johanson, D., Lovejoy, C., Kimbel, W., White, T., Ward, S., Bush, M., Latimer, B., & Coppens, Y. (1982). Morphology of the Pliocene partial hominid skeleton (A.L. 288-1) from the Hadar formation, Ethiopia American Journal of Physical Anthropology, 57, 403-451 DOI: 10.1002/ajpa.1330570403
A week and a half ago, Kibii and colleagues (2011) published reconstructions and re-analyses of two hips belonging to the 1.98 million-year old Australopithecus sediba. As with many fossil discoveries, these additions to the fossil record raise more questions than they answer. Unless the question was, “did A. sediba have a pelvis?” It did. Here’s a good summary from the paper itself:
Thus, Au. sediba is australopith-like in having a long superior pubic ramus and an anteriorly positioned and indistinctly developed iliac pillar…[and] Homo-like in having vertically oriented and sigmoid shaped iliac blades, more robust ilia, and a narrow tuberoacetabular sulcus…and the pubic body is upwardly rotated as in Homo. (p. 1410, emphases mine)
So far as I can tell, the main way the hips are ‘advanced’ toward a more human-like condition is that the iliac blades are more upright and sweep forward more than in earlier known hominid hips. Here’s the figure 2 from the paper (more sweet pics of the fossils are available here). NB that in both A. sediba hips much of the upper portions of the iliac blades are missing (reconstructed in white; this region is missing in lots of fossils), so it’s possible they were more flaring like the australopith in the center photo.
The authors’ bottom-line, take-home point is that the A. sediba pelvis has features traditionally associated with large-brained Homo – but belonged to a small-brained species (based solely on the ~430 cc MH1 endocast). They argue that this means that many of these unique pelvic features did not evolve in the context of birthing large-brained babies, as has often been thought. They state that these features are thus “most parsimoniously attributed to altered biomechanical demands on the pelvis in locomotion,” and suggest that this hypothetical locomotion was mostly bipedalism but with a good degree of climbing. Maybe, maybe not. This interpretation is consistent with the analysis of the A. sediba foot/ankle (Zipfel et al. 2011).
The weird mix of ancient (australopith-like) and newer (Homo-like) pelvic features in A. sediba really raises the question of how australopithecines moved around. More intriguing is that the A. sediba pelvis has differentHomo-like features than the ~1 million year old Busidima pelvis (Simpson et al. 2008), which has been attributed to Homo erectus (largely in aspects of the iliac blades). This raises the question of whether A. sediba is really pertinent to the origins of the genus Homo, and whether the Busidima pelvis belongs to Homo erectus or a late-surviving robust australopithecus (e.g. boisei, Ruff 2010).
Also interesting is that the subpubic angle (in the pic above, the upside-down “V” created by the pubic bones just above the red labels) is pretty low in MH2. This is curious because modern human males and females differ in how large this angle is – females tend to have a large angle which contributes to an enlarged birth canal, whereas males have a low angle like MH2. But MH2 is considered female based on skeletal and dental size. This raises the additional questions of whether human-like sexual dimorphism had not evolved in hominids prior to 1.9 million years ago, and whether the sex of MH2 was accurately described.
Finally, though the authors did a great job comparing this pelvis with those from other hominids, I think a major, more comprehensive comparative review of hominid pelves is in order. How does the older A. afarensis hip from Woranso (Haile-Selassie et al. 2010) inform australopithecine pelvic evolution? What about the possibly-contemporary-maybe-later hip from the nearby site of Drimolen (Gommery et al. 2002)? Given the subadult status of the MH1 individual, it would be interesting to compare with the WT 15000 Homo erectus fossils, or A. africanus subadults from Makapansgat, to examine the evolution of pelvic growth.
Lots of interesting questions arise from these fascinating new fossils. “The more you know,” right?
References Gommery, D. (2002). Description d’un bassin fragmentaire de Paranthropus robustus du site Plio-Pléistocène de Drimolen (Afrique du Sud)A fragmentary pelvis of Paranthropus robustus of the Plio-Pleistocene site of Drimolen (Republic of South Africa) Geobios, 35 (2), 265-281 DOI: 10.1016/S0016-6995(02)00022-0
Haile-Selassie Y, Latimer BM, Alene M, Deino AL, Gibert L, Melillo SM, Saylor BZ, Scott GR, & Lovejoy CO (2010). An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia. Proceedings of the National Academy of Sciences of the United States of America, 107 (27), 12121-6 PMID: 20566837
Kibii, J., Churchill, S., Schmid, P., Carlson, K., Reed, N., de Ruiter, D., & Berger, L. (2011). A Partial Pelvis of Australopithecus sediba Science, 333 (6048), 1407-1411 DOI: 10.1126/science.1202521
Ruff, C. (2010). Body size and body shape in early hominins – implications of the Gona Pelvis Journal of Human Evolution, 58 (2), 166-178 DOI: 10.1016/j.jhevol.2009.10.003
Simpson, S., Quade, J., Levin, N., Butler, R., Dupont-Nivet, G., Everett, M., & Semaw, S. (2008). A Female Homo erectus Pelvis from Gona, Ethiopia Science, 322 (5904), 1089-1092 DOI: 10.1126/science.1163592
Zipfel, B., DeSilva, J., Kidd, R., Carlson, K., Churchill, S., & Berger, L. (2011). The Foot and Ankle of Australopithecus sediba Science, 333 (6048), 1417-1420 DOI: 10.1126/science.1202703
National Geographic aired a special tonight about a recently-excavated child’s skeleton (they focused on the skull) from Grotte des Contrebandiers in Morocco, dated to around 108,000 years ago. So far as I know this material has not been fully published (aside from a brief blurb in Science).
The program presented work of archaeologists, paleontologists, reconstruction artists, taphonomists, and lots of other people, hoping to figure out who the kid was and such. All in all it was pretty cool, I’d recommend checking it out if you didn’t see it. Or again if you did see it.
While I think it was a great program and the researchers involved are doing a terrific job, I had two main notes: first, I wish they’d treated the topic of growth-n-development a little more. They noted that the child (5-6 years old possibly) looked really “modern” because of its flat face. But looking at it, it didn’t really have that flat of a face, especially for a child. They talked about how human-like (rather than Neandertal-like) the kid was, but they only compared it with adults – children tend to have relatively smaller faces and larger brain-cases than adults (right), so it’s no wonder it looked more like an adult human than the adult Neandertal from Amud (Israel) that they compared it with. It would’ve been great to see more comparisons with other late Pleistocene hominid kids, such as from Skhul/Qafzeh or La Quina. A future program, perhaps.
Second, they kept asking whether the kid was “a Homo sapien.” I know it’s counterintuitive for English-speakers, but “H. sapiens” is the singular and plural of humans’ scientific name. Silly, right, cuz it doesn’t even get paid twice as much. But you’ll have take that up with C. Linnaeus. I am a Homo sapiens. You are a Homo sapiens. Fifty people are a gaggle of Homo sapiens.
Anyway it was a cool show. Check it out!
Figure credit: Fig. 2 from Bogin. 2003. The human pattern of growth and development in paleontological perspective. In Patterns of Growth and Development in the Genus Homo, eds. Thompson JL, Krovitz GE and Nelson AJ. New York: Cambridge University Press: 15-44.