We just kicked off the Spring semester here at Vassar College, and so I’ve got some freshly-updated bio-anthro syllabi hot off the press. This semester, I’m doing my annual introductory class (Anth 120, “Human Origins”), a resurrected seminar (Anth 305: “Human Evo-Devo”), and a second stab at a new methods module (Anth 211: “Virtual Anthropology”).
Anth 120 is similar to previous versions, although this year I’ve taken out a reading/lecture on Paleolithic technology, replaced with articles scrutinizing evolutionary psychology. We’ll see how it goes.
The other two classes are greatly overhauled from previous versions. Anth 211, “Virtual Anthropology,” is my first contribution to a new curricular initiative here at Vassar, which are called “intensives.” Anth 211 is kind of a hybrid between a regular class and an independent study, giving students experience with computer-based, “virtual” methods used in biological anthropology and related fields. In the first half of the semester, students will get to try out some of these methods and see what kinds of research questions they’re used for. In the 2nd half of the term, students do their own Virtual Anthropology study drawing on the materials in my HEAD Lab, and then present a research poster at the end of the year. I debuted this intensive last Fall, and based on that experience I’m providing a bit more training and have more activities for students this Spring. If last semester’s projects are at all predictive, we should have some fun projects in store this year.
Anth 305 is a fossil-focused examination of the roles of growth and development in human evolution, and this year’s version is also highly modified from the last time I taught it over two years ago. In that first version, course content was patterned along the skeleton, e.g., one week looked at evolution and development of teeth, next week the spine, etc. Such a bauplan might work for building bodies, but it wasn’t the best for teaching. So this year, we’re spending the first few weeks on the fossil record of human evolution, getting acquainted with the curious characters of our deep past. From there, we go over skeletal / developmental biology, before delving into special evo-devo topics like “morphological integration” and “heterochrony” for the rest of the semester. We’ll also read lots of old, “classic” papers along the way.
Syllabi for these, and other classes, can be found on the teaching page of the site, if you want to learn more.
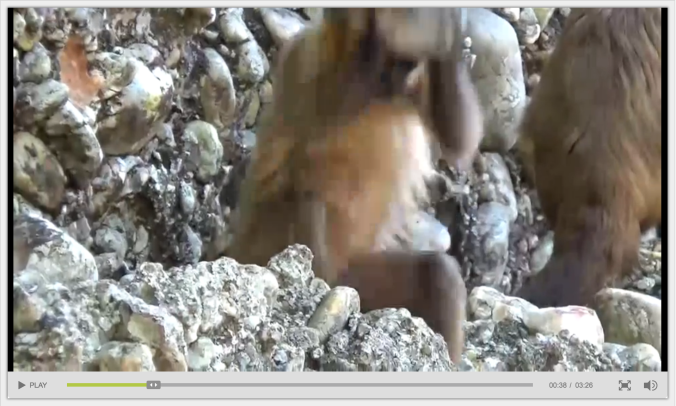
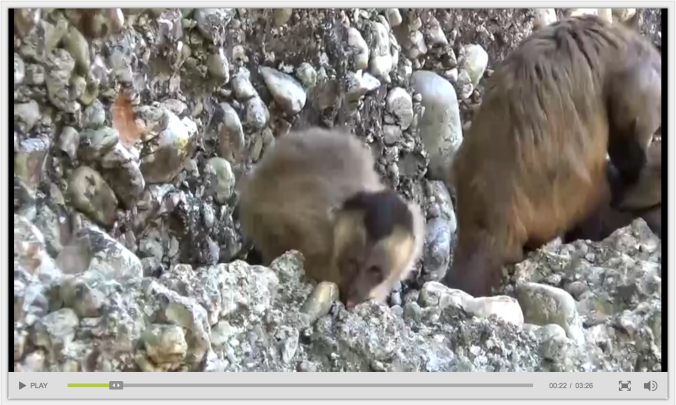
A month ago at ESHE and now online in Nature, Proffitt and colleagues describe stone-on-stone smashing behavior among wild bearded capuchin monkeys (Sapajus libidinosus). The online paper includes a great video documenting the action; here’s a screenshot:
Holding the rock with both hands just above head-level, the monkey prepares to crush its enemies. Which in this case are another rock stuck in a pile of more rocks.
In the fairly rare cases where non-human primates use stones, it’s for smashing nuts or something. But when these capuchins see a stone they don’t just see a smasher, they see a world of possibilities* – why use a rock to break a rock, when you could use it to break a heart? So this group of capuchins is unique in part because they’ve been documented to use stones for many purposes.
Now why on earth a monkey would use one rock to break another rock is anyone’s guess. In human evolution, the purpose was to break off small, sharp flakes that could be used to butcher animals or work plants. Proffitt et al. did observe small flakes being removed when capuchins pounded rocks, but the monkeys showed little interest in this debitage, other than using it to continue smashing stuff. More curiously, the monkeys frequently lick the rock after hammering at it:
Mmm, rocks.
Proffitt et al. venture that maybe these monkeys are doing this to ingest lichens or trace elements like silicon. This hypothesis merits further investigation, but what’s clear is that these monkeys’ lithics differ from the hominin archaeological record wherein the express purpose of breaking rocks is to make flakes.
What’s striking to me (pun intended) is the relative size of the rocks. These monkeys that weigh only 2-3 kg are lifting and smashing stones that weigh about half a kilogram on average. Because these stones are fairly large given the monkeys’ body size, they have to be lifted with two hands and brought down on a surface, a “passive hammer” technique. The earliest-known tools made by hominins, from the 3.3 million year old Lomekwi site in Kenya, are also pretty big. Weighing 3 kg on average but topping at 15 kg, these earliest tools would have required the same knapping technique as is used by these little monkeys (Harmand et al., 2015).
Left: Cover of Nature vol. 521 (7552). Right: Bearded capuchin letting a pebble know who’s boss (link).
Why the big stuff at first? Did the earliest hominin tool-makers lack the dexterity to make tools from the smaller rocks comprising the later Oldowan industry? These creative capuchins could lead to predictions about the hand/arm skeleton of the Lomekwian tool-makers (testable, of course, only with fortuitous fossil discoveries). Capuchins are noted for their manual dexterity (Truppa et al., 2016) and have a similar thumb-index finger ratio to humans and early hominins (Feix et al. 2015), although they differ from humans in the insertion of the opponens muscle and resultant mobility of the thumb (Aversi-Ferreira et al., 2014). Maybe these tech-smart monkeys can tell us more about the earliest human tool-makers’ bodies than their brains.
References
Aversi-Ferreira RA, Souto Maior R, Aziz A, Ziermann JM, Nishijo H, Tomaz C, Tavares MC, & Aversi-Ferreira TA (2014). Anatomical analysis of thumb opponency movement in the capuchin monkey (Sapajus sp). PloS one, 9 (2) PMID: 24498307
Feix T, Kivell TL, Pouydebat E, & Dollar AM (2015). Estimating thumb-index finger precision grip and manipulation potential in extant and fossil primates. Journal of the Royal Society, Interface, 12 (106) PMID: 25878134
Harmand S, Lewis JE, Feibel CS, Lepre CJ, Prat S, Lenoble A, Boës X, Quinn RL, Brenet M, Arroyo A, Taylor N, Clément S, Daver G, Brugal JP, Leakey L, Mortlock RA, Wright JD, Lokorodi S, Kirwa C, Kent DV, & Roche H (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521 (7552), 310-5 PMID: 25993961
Proffitt, T., Luncz, L., Falótico, T., Ottoni, E., de la Torre, I., & Haslam, M. (2016). Wild monkeys flake stone tools Nature DOI: 10.1038/nature20112
Truppa V, Spinozzi G, Laganà T, Piano Mortari E, & Sabbatini G (2016). Versatile grasping ability in power-grip actions by tufted capuchin monkeys (Sapajus spp.). American Journal of Physical Anthropology, 159 (1), 63-72 PMID: 26301957
*well, at least four uses given by Proffitt et al.: mating display, aggression, food-crushing, and digging.
Holy crap 2015 was a big year for fossils. And how fortuitous that 2016 begins on a Fossil Friday – let’s recap some of last year’s major discoveries.
Homo naledi
Some Homo naledi mandibles in order from least to most worn teeth.
The Homo naledi sample is a paleoanthropologist’s dream – a new member of the genus Homo with a unique combination of traits, countless remains belonging to at leasta dozen individuals from infant to old adult, representation of pretty much the entire skeleton, and a remarkable geological context indicative of intentional disposal of the dead (but certainly not homicide, grumble grumble grumble…). The end of 2015 saw the announcement and uproar (often quite sexist) over this amazing sample. You can expect to see more, positive things about this amazing animal in 2016.
We’ll be presenting a bunch about Homo naledi at this year’s AAPA meeting in Hotlanta. I for one will be discussing dental development at Dinaledi- here’s a teaser:
As long as we’re talking about the AAPA meetings, my colleague David Pappano and I are organizing a workshop, “Using the R Programming Language for Biological Anthropology.” Details to come!
Lemur graveyard
Homo naledi wasn’t the only miraculously copious primate sample announced in 2015. Early last year scientists also reported the discovery of an “Enormous underwater fossil graveyard,” containing fairly complete remains of probably hundreds of extinct lemurs and other animals. As with Homo naledi, such a large sample will reveal lots of critical information about the biology of these extinct species.
Australopithecus deyiremeda
Extended Figure 1h from Haile-Selassie et al. (2015), compared with Demirjian developmental stages 6-8 . While the M1 roots look like stage 8 (complete), M2 looks like stage 7 (incomplete).
We also got a new species of australopithecus last year. Australopithecus deyiremeda had fat mandibles, a relatively short face (possibly…), and smaller teeth than in contemporaneous A. afarensis. One tantalizing thing about this discovery is that we may finally be able to put a face to the mysterious foot from Burtele, since these fossils come from nearby sites of about the same geological age. Also intriguing is the possible evidence, based on published CT images (above), that A. deyiremeda had relatively advanced canine and delayed molar development, a pattern generally attributed to Homo and not other australopithecines (if this turns out to be the case, you heard it here first!).
Lomekwian stone tool industry
3D scan and geographical location of Lomekwian tools. From africanfossils.org.
Roughly contemporaneous with A. deyiremeda, Harmand et al. (2015) report the earliest known stone tools from the 3.3 million year old site of Lomekwi 3 in Kenya. These tools are a bit cruder and much older than the erstwhile oldest tools, the Oldowan from 2.6 million years ago. These Lomekwian tools, and possible evidence for animal butchery at the 3.4 million year old Dikika site in Ethiopia (McPherron et al. 2010; Thompson et al. 2015), point to an earlier origin of lithic technology. Fossils attributed to Kenyanthropus platyops are also found at other sites at Lomekwi. With hints at hominin diversity but no direct associations between fossils and tools at this time, a lingering question is who exactly was making and using the first stone tools.
Earliest Homo
The reconstructed Ledi Geraru mandible (top left), compared with Homo naledi (top right), A. deyiremeda (bottom left), and the Uraha early Homo mandible from Malawi (bottom right). Jaws are scaled to roughly the same length from the front to back teeth; the Uraha mandible does not have an erupted third molar whereas the others do and are fully adult.
Just as Sonia Harmand and colleagues pushed back the origins of technology, Brian Villmoare et al. pushed back the origins of the genus Homo, with a 2.7 million year old mandible from Ledi Geraru in Ethiopia. This fossil is only a few hundred thousand years younger than Australopithecus afarensis fossils from the nearby site of Hadar. But the overall anatomy of the Ledi Geraru jaw is quite distinct from A. afarensis, and is much more similar to later Homo fossils (see image above). Hopefully 2016 will reveal other parts of the skeleton of whatever species this jaw belongs to, which will be critical in helping explain how and why our ancestors diverged from the australopithecines. (note that we don’t yet have a date for Homo naledi – maybe these will turn out to be older?)
Early and later Homo
Left: modified figures 2-3 from Maddux et al. (2015). Right: modified figures 7 & 13 from Ward et al. (2015). Note that in the right plot, ER 5881 femur head diameter is smaller than all other Homo except BSN 49/P27.
The earlier hominin fossil record wasn’t the only part to be shaken up. A small molar (KNM-ER 51261) and a set of associated hip bones (KNM-ER 5881) extended the lower range of size variation in Middle and Early (respectively) Pleistocene Homo. It remains to be seen whether this is due to intraspecific variation, for example sex differences, or taxonomic diversity; my money would be on the former.
Left: Penghu 1 hemi-mandible (Chang et al. 2015: Fig. 3), viewed from the outside (top) and inside (bottom). Right: Manot 1 partial cranium (Hershkovitz et al. 2015: Fig. 2), viewed from the left (top) and back (bottom).
At the later end of the fossil human spectrum, researchers also announced an archaic looking mandible dredged up from the Taiwan Straits, and a more modern-looking brain case from Israel. The Penghu 1 mandible is likely under 200,000 years old, and suggests a late survival of archaic-looking humans in East Asia. Maybe this is a fossil Denisovan, who knows? What other human fossils are waiting to be discovered from murky depths?
The Manot 1 calvaria looks very similar to Upper Paleolithic European remains, but is about 20,000 years older. At the ESHE meetings, Israel Hershkovitz actually said the brain case compares well with the Shanidar Neandertals. So wait, is it modern or archaic? As is usually the case, with more fossils come more questions.
Crazy dinosaurs
Yi qi was bringing Skeksi back, and its upper limb had a wing-like shape not seen in any other dinosaur, bird or pterosaur. There were a number of other interesting non-human fossil announcements in 2015 (see here and here), proving yet again that evolution is far more creative than your favorite monster movie makers.
What a year – new species, new tool industries, new ranges of variation! 2015 was a great year to be a paleoanthropologist, and I’ll bet 2016 has just as much excitement in store.
References (in order of appearance)
Haile-Selassie, Y., Gibert, L., Melillo, S., Ryan, T., Alene, M., Deino, A., Levin, N., Scott, G., & Saylor, B. (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity Nature, 521 (7553), 483-488 DOI: 10.1038/nature14448
Harmand, S., Lewis, J., Feibel, C., Lepre, C., Prat, S., Lenoble, A., Boës, X., Quinn, R., Brenet, M., Arroyo, A., Taylor, N., Clément, S., Daver, G., Brugal, J., Leakey, L., Mortlock, R., Wright, J., Lokorodi, S., Kirwa, C., Kent, D., & Roche, H. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521 (7552), 310-315. DOI: 10.1038/nature14464
McPherron, S., Alemseged, Z., Marean, C., Wynn, J., Reed, D., Geraads, D., Bobe, R., & Béarat, H. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature, 466 (7308), 857-860. DOI: 10.1038/nature09248
Thompson, J., McPherron, S., Bobe, R., Reed, D., Barr, W., Wynn, J., Marean, C., Geraads, D., & Alemseged, Z. (2015). Taphonomy of fossils from the hominin-bearing deposits at Dikika, Ethiopia Journal of Human Evolution, 86, 112-135 DOI: 10.1016/j.jhevol.2015.06.013
Villmoare, B., Kimbel, W., Seyoum, C., Campisano, C., DiMaggio, E., Rowan, J., Braun, D., Arrowsmith, J., & Reed, K. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia Science, 347 (6228), 1352-1355 DOI: 10.1126/science.aaa1343
Maddux, S., Ward, C., Brown, F., Plavcan, J., & Manthi, F. (2015). A 750,000 year old hominin molar from the site of Nadung’a, West Turkana, Kenya Journal of Human Evolution, 80, 179-183 DOI: 10.1016/j.jhevol.2014.11.004
Ward, C., Feibel, C., Hammond, A., Leakey, L., Moffett, E., Plavcan, J., Skinner, M., Spoor, F., & Leakey, M. (2015). Associated ilium and femur from Koobi Fora, Kenya, and postcranial diversity in early Homo Journal of Human Evolution, 81, 48-67 DOI: 10.1016/j.jhevol.2015.01.005
Chang, C., Kaifu, Y., Takai, M., Kono, R., Grün, R., Matsu’ura, S., Kinsley, L., & Lin, L. (2015). The first archaic Homo from Taiwan Nature Communications, 6 DOI: 10.1038/ncomms7037
Hershkovitz, I., Marder, O., Ayalon, A., Bar-Matthews, M., Yasur, G., Boaretto, E., Caracuta, V., Alex, B., Frumkin, A., Goder-Goldberger, M., Gunz, P., Holloway, R., Latimer, B., Lavi, R., Matthews, A., Slon, V., Mayer, D., Berna, F., Bar-Oz, G., Yeshurun, R., May, H., Hans, M., Weber, G., & Barzilai, O. (2015). Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans Nature, 520 (7546), 216-219 DOI: 10.1038/nature14134
Matt Skinner and colleagues report in today’s Science an analysis of trabecular bone structure in the hand bones of humans, fossil hominins and living apes. Trabecular bone, the sponge-like network of bony lattices on the insides of many of your bones, adapts during life to better withstand the directions and amounts of force it experiences. This is a pretty great property of the skeleton: bone is organized in a way that helps withstand usual forces, and the spongy organization of trabeculae also keeps bones fairly lightweight. Win-win.
An X-ray of my foot. The individual foot bones are filled with narrow spicules (=trabeculae) of bone. Very often they have a directed, or non-random, orientation: in the calcaneus, for instance, they are oriented mostly from the heel to the ankle joint.
This adaptive nature of trabecular bone also means that we can learn a lot about how animals lived in the past when all they’ve left behind are scattered fossils. In the present case, Skinner and colleagues tested whether tool use leaves a ‘trabecular signature’ in hand bones, looking then for whether fossil hominins fit this signature. Their study design is beautifully simple but profoundly insightful: First, they compared humans and apes to see if the internal structure of their hand bones can be distinguished. Second, they tested whether these differences accord with theoretical predictions based on how these animals use their hands (humans manipulate objects, apes use hands for walking and climbing). Third, they determined whether fossil hand bones look more like either group.
Comparison of first metacarpals (the thumb bone in your palm) between a chimpanzee (left), three australopithecines, and a human (right). In each, the palm side is to the left and the wrist end of the bone (proximal) is down. Image by Tracy Kivell, and found here.
Looking at the image above, it’s difficult to spot trabecular differences between the specimens with the naked eye. But computer software can easily measure the density and distribution of trabecular bone from CT scans. With these tools, researchers found key differences between humans and apes consistent with the different ways they use their hands. Neandertals (humans in the past 100 thousand years or so) showed the human pattern, not unexpected since their bones look like ours and they used their hands to make tools and manipulate objects like we do.
What’s more interesting, though, is that the australopithecines, dating to between 1.8-3.0 million years ago, also show the human pattern. This is an important finding since the external anatomy of Australopithecus hand bones shows a mixture of human- and ape-like features, with unclear implications for how they used their hands. Their trabecular architecture, reflecting the forces their hands experienced in life, is consistent with tool use.
This is a very significant finding.Australopithecus africanus fossils from Sterkfontein aren’t associated with any stone tools; bone tools are known from Swartkrans, though it is unclear whether Australopithecus robustus or Early Homo from the site made/used these. In addition, in 2010 McPherron and colleagues reported on a possibly cut-marked animal bone from the 3.4 million year old site of Dikika in Ethiopia, where Australopithecus afarensis fossils but no tools are found. Skinner and colleagues’ results show that at the very least, South African Australopithecus species were using their hands like tool-makers and -users do.
This raises many fascinating questions – were australopithecines using stone tools, but we haven’t found them? Were they using tools made of other materials? What do the insides of Australopithecus afarensis metacarpals look like? What I like about this study is that it presents both compelling results, and raises further (testable) questions about both the nature of the earliest tools and our ability to detect their use from fossils.
I’m doing some reading on the study of craniofacial growth, I stumbled across this poignant quote from Melvin Moss, from a seminar on “New Techniques in Processing and Handling Growth Data”:
“This is very beautiful. It is neat, it is modern technology, and it is fast. I am just wondering very seriously about the biological validity of what we are doing with this machine.” (Moyers & Krogman, eds: p. 326)
Reference
Cranio-Facial Growth in Man. RE Moyers & WM Krogman, eds. 1967. New York: Pergamon Press.