The Fall semester here at Vassar kicks off next week, and so of course a new fossil discovery is published this week that threatens to upend my course plans and throw my syllabi into disarray. Haile-Selassie and colleagues report a very well-preserved hominin cranium, from the Woranso-Mille region of Ethiopia and dating to 3.8 million years ago. The new cranium shares features with Australopithecus anamensis, a species previously mainly known through jaws and teeth. The fossil is therefore really important since it puts a face to the species’ name, and it is the oldest relatively complete Australopithecus cranium known. When I showed a picture of the fossil to my wife, who is not a paleoanthropologist, all she said was that it looked like the face of a dog who got stung by a bee.
The new A. anamensis fossil MRD-VP-1 (left), and a dog that lost a fight with a bee. Fossil photo from the Smithsonian‘s coverage.
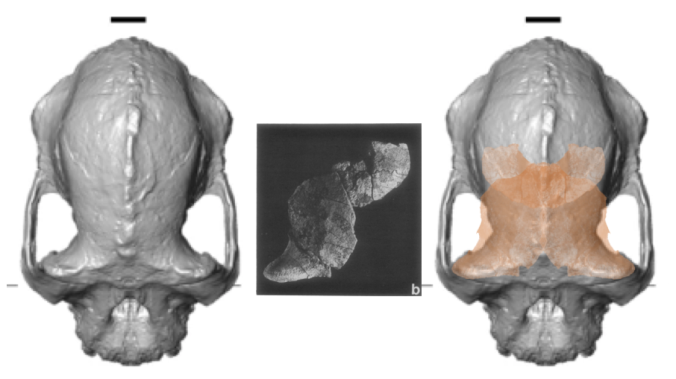
The big buzz in many news stories about the fossil (for example, Nature, ScienceNews, etc.) is that it rewrites an evolutionary relationship early in human history, with Australopithecus anamensis no longer the ancestor of A. afarensis, but rather the two being contemporaries. That idea is based on a 3.9 million year old frontal bone attributed to A. afarensis from a site called Belohdelie, also in Ethiopia (Asfaw, 1987): basically, the new A. anamensis cranium reveals a hominin with a narrow frontal region of the brain, which lived 100,000 later than A. afarensis with a relatively expanded frontal region:
Top views of the reconstructed A. anamensis cranium (left), and the Belohdelie frontal (center), and my crappy photoshopped overlay of Belohdelie on A. anamensis (right). Images not to scale.
The lede, “human evolutionary tree messier than thought,” is not terribly interesting or compelling since it seems to characterize most fossil discoveries over the past several years. And in this case I don’t know how well supported the argument is, since the trait in question (narrow frontal region of the braincase or “post-orbital constriction”) can vary dramatically within a single species. The image below is from the paper itself—compare the difference in “postorbital constriction index” (left graph) between the new A. anamensis cranium (MRD) and A. afarensis (in blue). Both sets of fossils fall within the range of chimpanzees (P. troglodytes), and note the great range of variation within gorillas (G. gorilla).
Part of Figure 3 from the paper by Haile-Selassie and colleagues. On the top is a view from above of fossil humans: Sahelanthropus tchadensis, Ardipithecus ramidus, the new A. anamensis, A. afarensis, and A. africanus. Below the graphs show how species differ in narrowing of the frontal (left) and length of the skull (right).
What I find most interesting about the new find is the great front-to-back length of the cranium—check out how long and narrow the brain-case is of the fossil compared with the later hominins to the right. This is an interesting similarity with the much earlier (6 million years ago) Sahelanthropus tchadensis, which is the left-most fossil in the figure. It makes me really curious to see the brain endocast of A. anamensis and the Sahelanthropus cranium—what was brain shape like for these ancient animals, and what does that mean for the earliest stages of human brain evolution? The Sahelanthropus endocast was meticulously reconstructed and analyzed by Thibaut Bienvenu (dissertation here), who found it to be somewhat like gorillas in overall shape (but with a more hominin-like placement of the foramen magnum). Of course, the outside of the skull doesn’t necessarily tell us what the underlying brain shape was like. Haile-Selassie and colleagues report that they made a virtual reconstruction of the A. anamensis endocast, so hopefully we’ll get to pick its brain soon.
Holy crap 2015 was a big year for fossils. And how fortuitous that 2016 begins on a Fossil Friday – let’s recap some of last year’s major discoveries.
Homo naledi
Some Homo naledi mandibles in order from least to most worn teeth.
The Homo naledi sample is a paleoanthropologist’s dream – a new member of the genus Homo with a unique combination of traits, countless remains belonging to at leasta dozen individuals from infant to old adult, representation of pretty much the entire skeleton, and a remarkable geological context indicative of intentional disposal of the dead (but certainly not homicide, grumble grumble grumble…). The end of 2015 saw the announcement and uproar (often quite sexist) over this amazing sample. You can expect to see more, positive things about this amazing animal in 2016.
We’ll be presenting a bunch about Homo naledi at this year’s AAPA meeting in Hotlanta. I for one will be discussing dental development at Dinaledi- here’s a teaser:
As long as we’re talking about the AAPA meetings, my colleague David Pappano and I are organizing a workshop, “Using the R Programming Language for Biological Anthropology.” Details to come!
Lemur graveyard
Homo naledi wasn’t the only miraculously copious primate sample announced in 2015. Early last year scientists also reported the discovery of an “Enormous underwater fossil graveyard,” containing fairly complete remains of probably hundreds of extinct lemurs and other animals. As with Homo naledi, such a large sample will reveal lots of critical information about the biology of these extinct species.
Australopithecus deyiremeda
Extended Figure 1h from Haile-Selassie et al. (2015), compared with Demirjian developmental stages 6-8 . While the M1 roots look like stage 8 (complete), M2 looks like stage 7 (incomplete).
We also got a new species of australopithecus last year. Australopithecus deyiremeda had fat mandibles, a relatively short face (possibly…), and smaller teeth than in contemporaneous A. afarensis. One tantalizing thing about this discovery is that we may finally be able to put a face to the mysterious foot from Burtele, since these fossils come from nearby sites of about the same geological age. Also intriguing is the possible evidence, based on published CT images (above), that A. deyiremeda had relatively advanced canine and delayed molar development, a pattern generally attributed to Homo and not other australopithecines (if this turns out to be the case, you heard it here first!).
Lomekwian stone tool industry
3D scan and geographical location of Lomekwian tools. From africanfossils.org.
Roughly contemporaneous with A. deyiremeda, Harmand et al. (2015) report the earliest known stone tools from the 3.3 million year old site of Lomekwi 3 in Kenya. These tools are a bit cruder and much older than the erstwhile oldest tools, the Oldowan from 2.6 million years ago. These Lomekwian tools, and possible evidence for animal butchery at the 3.4 million year old Dikika site in Ethiopia (McPherron et al. 2010; Thompson et al. 2015), point to an earlier origin of lithic technology. Fossils attributed to Kenyanthropus platyops are also found at other sites at Lomekwi. With hints at hominin diversity but no direct associations between fossils and tools at this time, a lingering question is who exactly was making and using the first stone tools.
Earliest Homo
The reconstructed Ledi Geraru mandible (top left), compared with Homo naledi (top right), A. deyiremeda (bottom left), and the Uraha early Homo mandible from Malawi (bottom right). Jaws are scaled to roughly the same length from the front to back teeth; the Uraha mandible does not have an erupted third molar whereas the others do and are fully adult.
Just as Sonia Harmand and colleagues pushed back the origins of technology, Brian Villmoare et al. pushed back the origins of the genus Homo, with a 2.7 million year old mandible from Ledi Geraru in Ethiopia. This fossil is only a few hundred thousand years younger than Australopithecus afarensis fossils from the nearby site of Hadar. But the overall anatomy of the Ledi Geraru jaw is quite distinct from A. afarensis, and is much more similar to later Homo fossils (see image above). Hopefully 2016 will reveal other parts of the skeleton of whatever species this jaw belongs to, which will be critical in helping explain how and why our ancestors diverged from the australopithecines. (note that we don’t yet have a date for Homo naledi – maybe these will turn out to be older?)
Early and later Homo
Left: modified figures 2-3 from Maddux et al. (2015). Right: modified figures 7 & 13 from Ward et al. (2015). Note that in the right plot, ER 5881 femur head diameter is smaller than all other Homo except BSN 49/P27.
The earlier hominin fossil record wasn’t the only part to be shaken up. A small molar (KNM-ER 51261) and a set of associated hip bones (KNM-ER 5881) extended the lower range of size variation in Middle and Early (respectively) Pleistocene Homo. It remains to be seen whether this is due to intraspecific variation, for example sex differences, or taxonomic diversity; my money would be on the former.
Left: Penghu 1 hemi-mandible (Chang et al. 2015: Fig. 3), viewed from the outside (top) and inside (bottom). Right: Manot 1 partial cranium (Hershkovitz et al. 2015: Fig. 2), viewed from the left (top) and back (bottom).
At the later end of the fossil human spectrum, researchers also announced an archaic looking mandible dredged up from the Taiwan Straits, and a more modern-looking brain case from Israel. The Penghu 1 mandible is likely under 200,000 years old, and suggests a late survival of archaic-looking humans in East Asia. Maybe this is a fossil Denisovan, who knows? What other human fossils are waiting to be discovered from murky depths?
The Manot 1 calvaria looks very similar to Upper Paleolithic European remains, but is about 20,000 years older. At the ESHE meetings, Israel Hershkovitz actually said the brain case compares well with the Shanidar Neandertals. So wait, is it modern or archaic? As is usually the case, with more fossils come more questions.
Crazy dinosaurs
Yi qi was bringing Skeksi back, and its upper limb had a wing-like shape not seen in any other dinosaur, bird or pterosaur. There were a number of other interesting non-human fossil announcements in 2015 (see here and here), proving yet again that evolution is far more creative than your favorite monster movie makers.
What a year – new species, new tool industries, new ranges of variation! 2015 was a great year to be a paleoanthropologist, and I’ll bet 2016 has just as much excitement in store.
References (in order of appearance)
Haile-Selassie, Y., Gibert, L., Melillo, S., Ryan, T., Alene, M., Deino, A., Levin, N., Scott, G., & Saylor, B. (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity Nature, 521 (7553), 483-488 DOI: 10.1038/nature14448
Harmand, S., Lewis, J., Feibel, C., Lepre, C., Prat, S., Lenoble, A., Boës, X., Quinn, R., Brenet, M., Arroyo, A., Taylor, N., Clément, S., Daver, G., Brugal, J., Leakey, L., Mortlock, R., Wright, J., Lokorodi, S., Kirwa, C., Kent, D., & Roche, H. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521 (7552), 310-315. DOI: 10.1038/nature14464
McPherron, S., Alemseged, Z., Marean, C., Wynn, J., Reed, D., Geraads, D., Bobe, R., & Béarat, H. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature, 466 (7308), 857-860. DOI: 10.1038/nature09248
Thompson, J., McPherron, S., Bobe, R., Reed, D., Barr, W., Wynn, J., Marean, C., Geraads, D., & Alemseged, Z. (2015). Taphonomy of fossils from the hominin-bearing deposits at Dikika, Ethiopia Journal of Human Evolution, 86, 112-135 DOI: 10.1016/j.jhevol.2015.06.013
Villmoare, B., Kimbel, W., Seyoum, C., Campisano, C., DiMaggio, E., Rowan, J., Braun, D., Arrowsmith, J., & Reed, K. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia Science, 347 (6228), 1352-1355 DOI: 10.1126/science.aaa1343
Maddux, S., Ward, C., Brown, F., Plavcan, J., & Manthi, F. (2015). A 750,000 year old hominin molar from the site of Nadung’a, West Turkana, Kenya Journal of Human Evolution, 80, 179-183 DOI: 10.1016/j.jhevol.2014.11.004
Ward, C., Feibel, C., Hammond, A., Leakey, L., Moffett, E., Plavcan, J., Skinner, M., Spoor, F., & Leakey, M. (2015). Associated ilium and femur from Koobi Fora, Kenya, and postcranial diversity in early Homo Journal of Human Evolution, 81, 48-67 DOI: 10.1016/j.jhevol.2015.01.005
Chang, C., Kaifu, Y., Takai, M., Kono, R., Grün, R., Matsu’ura, S., Kinsley, L., & Lin, L. (2015). The first archaic Homo from Taiwan Nature Communications, 6 DOI: 10.1038/ncomms7037
Hershkovitz, I., Marder, O., Ayalon, A., Bar-Matthews, M., Yasur, G., Boaretto, E., Caracuta, V., Alex, B., Frumkin, A., Goder-Goldberger, M., Gunz, P., Holloway, R., Latimer, B., Lavi, R., Matthews, A., Slon, V., Mayer, D., Berna, F., Bar-Oz, G., Yeshurun, R., May, H., Hans, M., Weber, G., & Barzilai, O. (2015). Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans Nature, 520 (7546), 216-219 DOI: 10.1038/nature14134
Last November I reported on recently described Australopithecus cf. afarensis craniodental remains from the site of Woranso Mille in Ethiopia. These fossils are significant in part because they date to around 3.6 million years ago; most of the postcranial evidence for A. afarensis comes from Hadar (~3.4 – 2.9 million years) or Maka (~3.5 million years). It is pretty awesome, then, that Yohannes Haile-Selassie and colleagues (2010a) have just reported on a partial skeleton from Woranso-Mille.
The specimen is given the catalog number KSD-VP-1/1 (right, from Nature), and the nickname Kadanuumuu, meaning “Big Man” in the language of the Afar people who live in the region where the fossils were discovered. Here, I’ll be focusing on the scapula.
Researchers have debated about what the scapular form of A. afarensis means functionally – how could, and did, these creatures use their shoulders? The scapula of AL 288 (the famous “Lucy”) preserves part of the glenoid fossa (shoulder socket) and only a little of the surrounding bone including the scapular spine (below). It has been argued that the angle between the glenoid fossa and the lateral border is more similar to modern apes than to humans. That is, the shoulder socket may have been oriented more upward, like in modern apes, compared to humans whose socket faces more to the side. The implication is that A. afarensis may have been preferentially exploiting arboreal environments.
Left: AL 288 scapular fragment. The glenoid fossa is the hollow that faces to the right, the lateral border is at the bottom paralleling the label “AL 288-1L.” The scapular spine is preserved only at the base, it is the small uprising of bone just to the left of the glenoid fossa. From Haile-Selassie et al. 2010b, Fig. S21.
Similarly, a juvenile afarensis skeleton from the Ethiopian site of Dikika (Alemseged et al. 2006), dating to around 3.4 million years ago, also suggested an ape-like shoulder for this extinct human ancestor. Principal components analysis of several measurements from the Dikika scapula showed it to be very similar to gorillas of comparable age, in terms of overall shape and proportions.
So from these two scapulae, one belonging to a very small-bodied female, the other from a small ~3-year-old possible female, we get the picture that A. afarensis had a fairly ape-like (i.e. arboreal) shoulder orientation, and may not have had independent movement of the head and trunk that we modern humans enjoy. Nevertheless, it is still unclear whether this means that the afarensis scapula functioned like that of an ape, and hence its shape, or whether the similarity in shape is a ‘hold-over’ from having an arboreal ancestor. I will say, I think one very telling feature noticeable in even the fragmentary AL 288 is the relative position and orientation of the scapular spine. Note that in the apes (the two juveniles scapulae on the right of the diagram to the left), the scapular spine roughly parallels the lateral border, and as a result, the flat areas above and below the spine are roughly equal in size. The above area houses the supraspinatus muscle, a rotator cuff muscle that acts largely in elevating the arm above the head and stabilizing the shoulder joint. In humans and afarensis, in contrast, the lower (insfraspinous) fossa is fairly large compared to the upper (supraspinous) fossa. Thus, the argument can be made that in humans and hominids, less power is needed to raise the arms over the head, or that humans and hominids have a greater reliance on the infraspinatus muscle for bringing the arm down toward the body and stabilizing the shoulder joint.
Now, KSD-VP-1 provides a remarkably complete scapula of an adult afarensis (right). In contrast to the specimens described above, KSD-VP-1 is very human-like. To the naked eye, and as borne out by principal components analysis of scapular angles, this thing is very human-like.
Now the question is, why does the morphology of this new specimen seem at odds with Lucy and Dikika? Part of the answer could be scaling – indeed, the authors note that the orientation of the glenoid relative to the lateral border (more specifically the scapular bar) in AL 288 can be found in modern humans of small size.
But that still does not answer the question of why the complete adult afarensis scapula is like adult humans, whereas the child afarensis is like young gorillas. The authors posit that perhaps it is due to Dikika’s fairly large supraspinous fossa. They also suggest that the measurements used in Alemseged et al’s study could not capture functional and discriminatory information about scapula shape. Nevertheless, a simple visual comparison the Dikika and KSD (x-ray…) scapulae reveals them to look fairly different, i.e. Dikika is relatively broader side-to-side.
Could ontogeny explain the differences between the child and adult afarensis? In a study of scapular growth and development in living primates, Young (2008) found childhood growth does not appear to explain adult shape variation. That is to say, most aspects of species-specific morphology are present in subadult scapulae. Rather, most variation in scapular shape among modern primates appears to be due to functional differences: climbers’ scapulae differ consistently from quadrupeds’. So what does that imply? That at 3.59 million years, adult male A. afarensis were not using their shoulders for arboreal activities, but at 3.4 million years ago, subadults were? Maybe this is just normal intraspecific variation? Maybe the ontogeny of the scapulae needs to be examined further?
I have to say I agree with Haile-Selassie et al. (2010a) here, that differences in the statistical analyses between the current study and that of Alemseged et al. (2006) may be partly responsible for the different interpretations of A. afarensis scapular morphology. Still, visual inspection of pictures of the fossils suggests to me that even if the principal components analyses were carried out using the same variables (Alemseged et al. used linear measurements, H-S et al. used angles), Dikika might seem gorilla-like, KSD still human-like; Nota bene that principal components analysis is not actually a test in itself, but rather an exploratory statistical technique. As such, it will never really “tell” how a bone was used. Still, I think this does raise an important issue about scapular function and ontogeny in hominoids.
References
Alemseged Z, Spoor F, Kimble WH, Bobe R, Geraads D, Reed D, and Wynn JG. 2006. A juvenile early hominin skeleton from Dikika, Ethiopia. Nature443: 296-301.
Haile-Selassie Y, Latimer BM, Alene M, Deino AL, Gibert L, Melillo, Saylor BZ, Scott GR, and Lovejoy CO. 2010a. An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia. Proceedings of the National Academy of Sciences, USA,in press.
Haile-Selassie et al. 2010b. Supplementary Online Material to 2010a.
Young NM. 2008. A Comparison of the Ontogeny of Shape Variation in the Anthropoid Scapula: Functional and Phylogenetic Signal. American Journal of Physical Anthropology136: 247-264.
Recently discovered Australopithecus fossils from the Ethiopian site of Woranso-Mille help fill a gap between parts of the early hominin fossil record (Haile-Selassie et al, in press).The fossils date to between 3.8-3.6 million years ago (Ma), and consist of several teeth and a jaw fragment. These specimens show a number of features that are intermediate in morphology between the earlier Au. anamensis (4.2-3.9 Ma) and later Au. afarensis from Laetoli (~3.7-3.5 Ma).As a result, the Woranso fossils lend support to the hypothesis that Au. anamensis and Au. afarensis represent a single evolving species (i.e. Kimbel et al. 2006).
I think this is exciting for two reasons.First, the fossils bridge the morphological gap between the older anamensis and younger afarensis fossils.As a result, we get to ‘see’ anagenetic evolution—changes within a single lineage.One topic in evolutionary biology is about the mode and tempo of evolution: are species fairly constant, then evolve into multiple ‘daughter’ species (“punctuated equilibrium”); or does evolutionary change tend to occur more within individual lineages (“anagenesis”)?Obviously neither is mutually exclusive, rather evolution is probably best characterized variously by both processes.Still, in the world of paleoanthropology, where many researchers argue for rapid and constant species turnover within the human lineage, it is cool to see a convincing argument for anagenesis.However, this ignores the meager (but intriguing) K. platyops material (Leakey et al. 2001), dating to around 3.5 Ma, possibly indicating the proliferation of at least two hominin species shortly after 4 Ma.
Second, the morphological intermediacy of the Woranso fossils allow a look at the patterns of evolutionary change within the anamensis–afarensis lineage.The authors note that the teeth of the Woranso hominins are generally more similar to anamensis, but have some derived characters of the later afarensis teeth. If we truly have a glimpse of dental evolution within a single lineage, we can ask questions about the evolution and development (“Evo-Devo”) of teeth. Are changes in these teeth correlated in a way that could be predicted by certain developmental models? Or is selection acting independently on various tooth traits?
References
Haile-Selassie Y, Saylor BZ, Deino A, Alene M, and Latimer BM. New hominid fossils from Woranso-Mille (Central Afar, Ethiopia) and Taxonomy of Early Australopithecus. American Journal of Physical Anthropology, in press.
Kimbel WH, Lockwood CA, Ward CV, Leakey MG, Rak Y, and Johanson DC. 2006. Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record. Journal of Human Evolution51: 134-152.
Leakey MG, Spoor F, Brown FH, Gathogo PN, Kiarie C, Leakey LN, and McDougall I. 2001. New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature410: 433-440.