Sorry I’m a bit late on this one, and that I’ve fallen behind on keeping the blog updated. I’ve been scrambling to make all the observations on, and collect all the data from, these
Australopithecus robustus mandibles in a short time. As my advisor likes to remind me, everything always takes 3x longer than you initially anticipate, and this is certainly true of my work here. Yesterday (the actual Fossil Friday), in fact, I probably spent only 30 min with these fossils. Instead, I accompanied Lee Berger and John Hawks on a trip to Malapa – the site that recently yielded fossils of the mysterious
Australopithecus sediba – and other sites in the area. To get there, I rented a car and drove on the wrong side of the road for the first time – it was a trippy trip, every time I got in the car I reached to my left for a phantom seat belt, and kept searching for the gear-shift my mind thought was in the door. Nuttiness.
Anyway, I have two thoughts for this edition of eFfing Fossil Friday. First point, related to the great tour from Dr. Berger, is that a ton of hominid fossils are lying in wait for us to re-expose them to the light of day. In South Africa, the classic Plio-Pleistocene sites have been Makapansgat (A. africanus), Sterkfontein (A. africanus) and Swartkrans (A. robustus and early Homo). These sites have variously been worked since the early 20th century. Since then, a number of other hominid-bearing sites – largely in the same area as Sterkfontein and Swartkrans – have been discovered: Gladysvale, Gondolin, Drimolen, and most recently Malapa. Yet still a metric-tonne of work is still being done on the more classic sites (except maybe Makapansgat?).
View of the valley, Malapa is somewhere in the background, I think the green patch of trees near the center, just before the big hill-shadow (?).
But these sites are just the tip of a fossiliferous iceberg. A few years ago when I was working here I accompanied some other researchers on a survey for more fossil sites in the area. What I learned then is that if you look across the Sterkfontein valley in the winter, the dessicated grassland is pimpled with the occasional patch of green trees – these small verdant isles are the tells of underlying cave systems (the caves contain water that plants will cut throats for). What was driven home yesterday at Malapa and other sites Dr. Berger showed us, is that these caves are all over the place, many fossil treasure-troves. What’s more, the A. sediba discovery (and the massive hominid molars from Gondolin) points to the idea that we are only beginning to understand what hominid life was like in the past. There is a rich prehistory still waiting to be discovered in South Africa, and undoubtedly also the rest of the African continent. Human paleontological work is far from exhausted. Let us usher in a Renaissance of field Paleoanthropology!
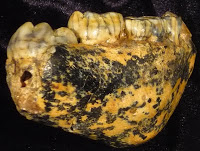
My next thought is that the process of fossilization can make the fossil-memories of past life quite beautiful. Now, in life the enamel of teeth is white-ish (yellow/brown is also not uncommon), and bone is this off-white/yellowish color. But during the process of fossilization, the original minerals used to make the bone (and less commonly teeth) are replaced by those in the surrounding soil. Often these minerals gussy up the fossils in neat new ways – manganese for example tends to make bone/tooth black.
Check out SK 61, an infant/child Australopithecus robustus. After fossilization, this thing takes on a designer, tortoise-shell coloration (left, above). SK 12, an older adult A. robustus (right, above), is another good example: some subterranean joker has drawn a smiley face beneath his left premolar (circled). So while we are often left with a meager fossil record, at least the fragments we get are voluptuously variegated.









{kind=link}
{kind=link}
{kind=link}