For the first time in many years, I’m offering a new advanced undergrad seminar here at Vassar. When I arrived here 8 years ago, I was mainly thinking about Homo naledi and ontogeny, so those were the foci of my seminars. But my research has begun looking more at brain evolution and especially the evidence from fossil endocasts, and there is a lot of literature I need to catch up on.
So I’ve invited students along for this brainstorm, using the question “Is the human brain special?” as a starting point to learn about how the beautifully congealed soup sloshing around inside our skull makes us such quirky animals. In the first half of the semester we’ll read up on brain anatomy and structure, and students will use some of the fossil endocast data I’ve accrued over the years to learn more about a given brain region and extinct hominin. In the second half of the semester we’ll read about the brains, behavior, and endocast fossils of very distant relatives — invertebrates, birds, whales, and dogs — that have been celebrated for their own ‘advanced intelligence.’ We’ll also read about how the evolution of our brains may have predisposed us to certain conditions like addiction and Alzheimer’s, and how brain science has been exploited toward racist and sexist ends (increasingly relevant in America today, sadly).
It will be a lot of work (I’m a very slow, distractible reader) but I’m excited to delve into this literature and see what insights our super sharp students here at Vassar come up with in discussions and projects. The course syllabus (ANTH 323) is available on my Teaching page — I’d be keen to hear suggestions for readings and assignments from folks who know more about brains than I do!
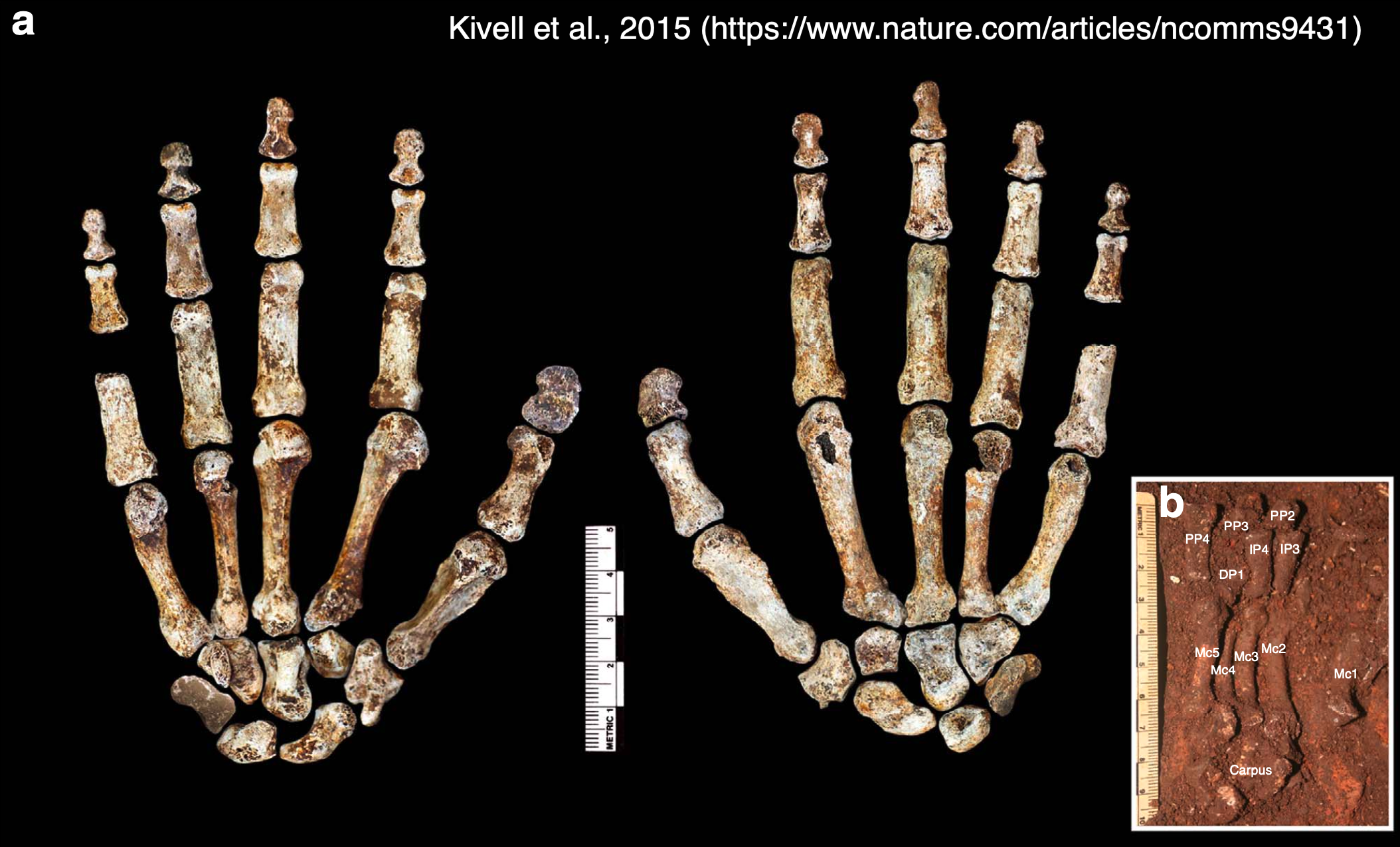
Homo naledi is one of my favorite extinct humans, in part because its impressive fossil record provides rare insights into patterns and process of growth and development. When researchers began recovering naledi fossils from Rising Star Cave 10 years ago, one of the coolest finds was this nearly complete hand skeleton. The individual bones were still articulated practically as they were in life so we know which bones belong to which fingers, allowing us grasp how dextrous this ancient human was. And since finger proportions are established before birth during embryonic development, we can see if Homo naledi bodies were assembled in ways more like us or other apes.
The “Hand 1” skeleton of Homo naledi, adapted from a figure by Kivell and colleagues (2015). Left shows the palm-side view while the middle shows the back of the hand. The inset (b) shows many of the palm and finger bones as they were found in situ in Rising Star Cave.
In a paper hot off the press (here), I teamed up with Dr. Tracy Kivell to analyze finger lengths of Homo naledi from the perspective of developmental biology. On the one hand, repeating structures such as teeth or the bones of a finger must be coordinated in their development, and scientists way smarter than me have come up with mathematical models predicting the relative sizes of these structures (for instance, teeth, digits, and more). On the other hand, the relative lengths of the second and fourth digits (pointer and ring fingers, respectively) are influenced by exposure to sex hormones during a narrow window in embryonic development: this ‘digit ratio’ tends to differ between mammalian males and females, and between primate species with different social systems.
So, Tracy and I examined the lengths of the three bones within the second digit (PP2, IP2, DP2) and of the first segment of the second and fourth digits (2P:4P) in Homo naledi, compared to published data for living and fossil primates (here and here). What did we find out?
Summary of our paper showing the finger segments analyzed (left), and graphs of the main results (right). The position of Homo naledi is highlighted by the blue star in each graph.
The first graph above compares the relative length of the first and last segments of the pointer finger across humans, apes, and fossil species. The dashed line shows where the data points are predicted to fall based on a theoretical model of development. There is a general separation between humans and the apes reflecting the fact that humans have a relatively long distal segment, which is important for precise grips when manipulating small objects. Fossil apes from millions of years ago and the 4.4 million year old hominin Ardipithecus are more like apes, while Homo naledi and more recent hominins are more like modern humans. Because both humans and apes fall close to the model predictions, this means the theoretical model does a good job of explaining how fingers develop. Because humans and apes differ from one another, this suggests a subtle ‘tweak’ to embryonic development may underlie the evolution of a precision grip in the human lineage, and that it occurred between the appearance of Ardipithecus and Homo.
The second graph compares the ‘digit ratio’ of the pointer and ring fingers from a handful of fossils with published ratios for humans and the other apes. Importantly, the digit ratio is high in gibbons (Hylobates) which usually form monogamous pair bonds, while the great apes (Pongo, Gorilla, Pan) are characterized by greater aggression and mating competition and have correspondingly lower digit ratios. Ever the bad primates, humans fall in between these two extremes. Most fossil apes and hominins have digit ratios within the range of overlap between the ape and human ratios, but Homo naledi has the highest ratio of all fossil hominins known, just above the human average. It has previously been suggested that humans’ higher ratio compared to earlier hominins may result from natural selection favoring less aggression and more cooperation recently in our evolution. If we can really extrapolate from digit proportions to behavior, this could mean Homo naledi was also less aggressive. This is consistent with the absence of healed skull fractures in the vast cranial sample (such skull injuries are common in much of the rest of the human fossil record).
You can see the amazing articulated Homo naledi hand skeleton for yourself on Morphosource. Its completeness reveals how handy Homo naledi was 300,000 years ago, and it can even shed light on the evolution of growth and development (and possibly social behavior) in the human lineage.
I’m working on a project analyzing infant remains of Homo naledi, a species of human that lived in South Africa around 300,000 years ago. In order to paint a full picture of infancy in this species, we need to estimate how big (or small) naledi newborns were. But without fossil neonates that could provide direct evidence of body size at birth, this is a tricky task.
Ideally, we could simply use adult body size estimates for Homo naledi to predict its body size at birth, using the scaling relationship in other primates as a guide. For example, using an average adult body size of 44 kg for Homo naledi (Garvin et al., 2017) yields an estimated newborn size of around 1.5 kg, based on published primate dataset (Barton and Cappellini, 2011). But this approach necessarily overlooks variation within each species, not to mention variation and uncertainty in Homo naledi adult size. In addition, the 95% prediction interval for this estimate ranges from under 1 kg (smaller than an average baboon baby) to almost as large as a human neonate.
Primate body size scaling (Barton & Cappellini, 2011). The black line is the regression for catarrhines (purple squares and blue circles), and the shaded grey area is the 95% prediction interval for newborns at a given adult catarrhine size.
And this gets at the other issue with the regression-based approach to estimating newborn body size in fossil hominins: humans are bad at being primates in some ways, as illustrated here by the fact that we don’t fit the newborn-adult body size relationship that characterizes other catarrhines (apes and monkeys of Africa and Eurasia).
Humans give birth to collosal kids. In contrast, gorillas are the largest living primates as adults, but their newborns are only a little over half the size of human neonates. Why do we have such giant babies? The most proximate reason is that humans are born with adult-ape-sized brains and quite a bit of baby fat as far as mammals go (Kuzawa, 1998). This tells us how babies are big, but it still begs the ultimate question of why—an enduring puzzle that you may have read about in the New York Times last week.
In order to land on a reasonable estimate of newborn body size in extinct humans, we need to figure out when evolution blew up the kid. Unfortunately, the only fossil hominin neonates are two Neandertals from France and Russia dating to under 100,000 years ago—pretty remarkable, but they don’t necessarily tell us about earlier species like Homo naledi.
My colleague Jerry Desilva (2011) worked out a potential solution to this conundrum. He argued that one could work from adult brain size to newborn body size through the following steps. First, in contrast to newborn-adult body size scaling, humans are good catarrhines when it comes to newborn-adult brain size scaling. This means that we can reasonably estimate newborn brain size based on adult brain sizes, which are aplenty in the human fossil record. Second, humans and many other primate newborns have brains roughly 12% of their overall body mass, while the great ape newborns stand out with brains around 10% of their adult size. Putting these two pieces together, one could estimate newborn body size: Adult brain ➡️ newborn brain ➡️ 10–12% newborn body size
DeSilva showed that regardless of whether you use an ape or human model of newborn brain/body size, hominin babies from Australopithecus afarensis 3 million years ago onward were probably large relative to maternal body size, estimated independently using skeletal remains. It’s a bit of a tortuous approach to estimating body size at birth, but the assumptions are reasonable and it’s probably the best way to figure out this important life history variable given the fossil evidence. What does this mean for Homo naledi?
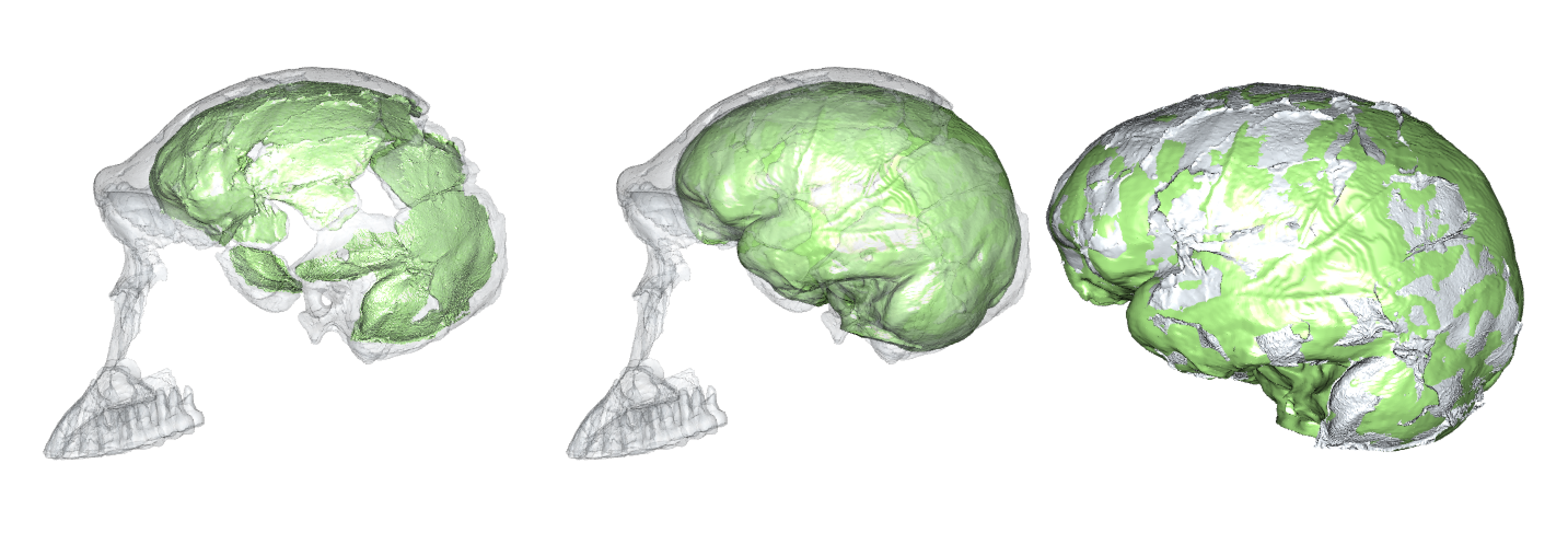
Virtual reconstruction of brain size and shape of the Homo naledi cranium “Neo” (work in progress). At 610 cm3, this is the largest and most complete Homo naledi endocast.
There are a few reliable adult brain size estimates for naledi, ranging from 465–610 cm3 (Berger et al., 2015; Garvin et al., 2017; Hawks et al., 2017), which based on catarrhine scaling would predict newborn brain size of around 170–210 cm3 (DeSilva and Lesnik, 2008). These brain sizes would then predict newborn body sizes of around 1.4–2.1 kg: the smol estimate is based on the smallest naledi adult brain size and a human model of newborn brain/body size; the chonk estimate is based on the largest naledi brain size and an ape brain/body model (pinkish stars in the boxplot below, left).
Boxplots of newborn body size in great apes. Gorilla, Chimpanzee, and Bonobo data from the Primate Aging Database(Kemnitz, 2019).
So, did Homo naledi have big babies? On the one hand, no: these 1.4–2.1 kg naledi newborns are outside the human range, and within the range of living great apes.
On the other hand, maybe Homo naledi babies were relatively large, though this depends on the size of Homo naledi adults. Recall from earlier that Garvin and colleagues arrived at an average estimated adult size of 44.2 kg. But this is an average of estimates for 20 separate naledi fossils, and each of these estimates has its own range of uncertainty. Garvin and team reported that the extremes of the prediction intervals for these estimates ranged from 28–62 kg. The second boxplot above shows newborn size relative to the adult average (sexes combined) for each species: for naledi, the six labels compare the smol and large newborn sizes (1.4 and 2.1 kg) with the adult average and extremes (28, 44, and 62 kg). Assuming the ‘true’ naledi sizes are somewhere in the middle of the range of estimates, naledi likely gave birth to babies 3–5% of adult body size, somewhat ‘intermediate’ between chimpanzees and humans (and bonobos…?) and similar to what DeSilva found for other hominins.
This is just a preliminary look at infancy in Homo naledi. There is a lot of uncertainty in these size estimates, but we should still be able to make some interesting inferences about growth and life history in our extinct evolutionary cousin.
REFERENCES
Barton, R. A., & Capellini, I. (2011). Maternal investment, life histories, and the costs of brain growth in mammals. Proceedings of the National Academy of Sciences, 108(15), 6169–6174. https://doi.org/10.1073/pnas.1019140108
Berger, L. R., Hawks, J., de Ruiter, D. J., Churchill, S. E., Schmid, P., Delezene, L. K., … Zipfel, B. (2015). Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. ELife, 4, e09560. https://doi.org/10.7554/eLife.09560
DeSilva, J. M. (2011). A shift toward birthing relatively large infants early in human evolution. Proceedings of the National Academy of Sciences, 108(3), 1022–1027. https://doi.org/10.1073/pnas.1003865108
DeSilva, J. M., & Lesnik, J. J. (2008). Brain size at birth throughout human evolution: A new method for estimating neonatal brain size in hominins. Journal of Human Evolution, 55(6), 1064–1074. https://doi.org/10.1016/j.jhevol.2008.07.008
Garvin, H. M., Elliott, M. C., Delezene, L. K., Hawks, J., Churchill, S. E., Berger, L. R., & Holliday, T. W. (2017). Body size, brain size, and sexual dimorphism in Homo naledi from the Dinaledi Chamber. Journal of Human Evolution, 111, 119–138. https://doi.org/10.1016/j.jhevol.2017.06.010
Hawks, J., Elliott, M., Schmid, P., Churchill, S. E., Ruiter, D. J. de, Roberts, E. M., … Berger, L. R. (2017). New fossil remains of Homo naledi from the Lesedi Chamber, South Africa. ELife, 6, e24232. https://doi.org/10.7554/eLife.24232
Kuzawa, C. W. (1998). Adipose tissue in human infancy and childhood: An evolutionary perspective. American Journal of Physical Anthropology, 107(S27), 177–209. https://doi.org/10.1002/(SICI)1096-8644(1998)107:27+<177::AID-AJPA7>3.0.CO;2-B
As we’re wrapping up what may be the worst year in recent global memory, especially geopolitically, let’s take a moment to review some more positive things that came up at Lawnchair in 2016.
Headed home
Alternate subtitle: Go West
This was a quiet year on the blog, with only 18 posts compared with the roughly thirty per year in 2014-2015. The major reason for the silence was that I moved from Kazakhstan back to the US to join the Anthropology Department at Vassar College in New York. With all the movement there was less time to blog. Much of the second half of 2016 was spent setting up the Biological Anthropology Lab at Vassar, which will focus on “virtual” anthropology, including 3D surface scanning…
Cast of early Homo cranium KNM-ER 1470 and 3D surface scan made in the lab using an Artec Spider.
… and 3D printing.
A gibbon endocast, created from a CT scan using Avizo software and printed on a Zortrax M200.
This first semester stateside I reworked my ‘Intro to Bio Anthro’ and ‘Race’ courses, which I think went pretty well being presented to an American audience for the first time. The latter class examines human biological variation, situating empirical observations in modern and historical social contexts. This is an especially important class today as 2016 saw a rise in nationalist and racist movements across the globe. Just yesterday Sarah Zhang published an essay in The Atlantic titled, “Will the Alt-right peddle a new kind of racist genetics?” It’s a great read, and I’m pleased to say that in the Race class this semester, we addressed all of the various social and scientific issues that came up in that piece. Admittedly though, I’m dismayed that this scary question has to be raised at this point in time, but it’s important for scholars to address and publicize given our society’s tragically short and selective memory.
So the first semester went well, and next semester I’ll be teaching a seminar focused on Homo naledi and a mid-level course on the prehistory of Central Asia. The Homo naledi class will be lots of fun, as we’ll used 3D printouts of H. naledi and other hominin species to address questions in human evolution. The Central Asia class will be good prep for when I return to Kazakhstan next summer to continue the hunt for human fossils in the country.
Osteology is still everywhere
A recurring segment over the years has been “Osteology Everywhere,” in which I recount how something I’ve seen out and about reminds me of a certain bone or fossil. Five of the blog 18 posts this year were OAs, and four of these were fossiliferous: I saw …
And a Homo erectus cranium on a Bangkok sidewalk. As I’m teaching a fossil-focused seminar next semester, OA will probably become increasingly about fossils, and I’ll probably get my students involved in the fun as well.
New discoveries and enduring questions
The most-read post on the blog this year was about the recovery of the oldest human Nuclear DNA, from the 450,000 year old Sima de los Huesos fossils. My 2013 prediction that nuclear DNA would conflict with mtDNA by showing these hominins to be closer to Neandertals than Denisovans was shown to be correct.
These results are significant in part because they demonstrate one way that new insights can be gained from fossils that have been known for years. But more intriguingly, the ability of researchers to extract DNA from exceedingly old fossils suggests that this is only the tip of the iceberg.
The comparison between monkey-made and anthropogenic stone tools drives home the now dated fact that humans aren’t the only rock-modifiers. But the significance for the evolution of human tool use is less clear cut – what are the parallels (if any) in the motivation and modification of rocks between hominins and capuchins, who haven’t shared a common ancestor for tens of millions of years? I’m sure we’ll hear more on that in the coming years.
In the case of whether Neandertal brain development is like that of humans, I pointed out that new study’s results differ from previous research probably because of differences samples and methods. The only way to reconcile this issue is for the two teams of researchers, one based in Zurich and the other in Leipzig, to come together or for a third party to try their hand at the analysis. Maybe we’ll see this in 2017, maybe not.
There were other cool things in 2016 that I just didn’t get around to writing about, such as the publication of new Laetoli footprints with accompanying free 3D scans, new papers on Homo naledi that are in press in the Journal of Human Evolution, and new analysis of old Lucy (Australopithecus afarensis) fossils suggesting that she spent a lifetime climbing trees but may have sucked at it. But here’s hoping that 2017 tops 2016, on the blog, in the fossil record, and basically on Earth in general.
The American winter holiday season is steeped in special spices, such as nutmeg, cloves, cinnamon, and whatever the hell pumpkin spice is. I guess as part of the never-ending War on Christmas, each year this sensory and commercial immersion begins earlier and earlier. Since these have become old news, I’d pretty much forgotten about the seasonal spicecapade until just the other day. In prep for minor holiday gluttony, I was grinding fresh nutmeg when I made a startling discovery. Nutmeg is not just the fragrant fruit of the Myristica fragrans tree. No, there’s something far more sinister in this holiday staple.
Merely nutmeg?
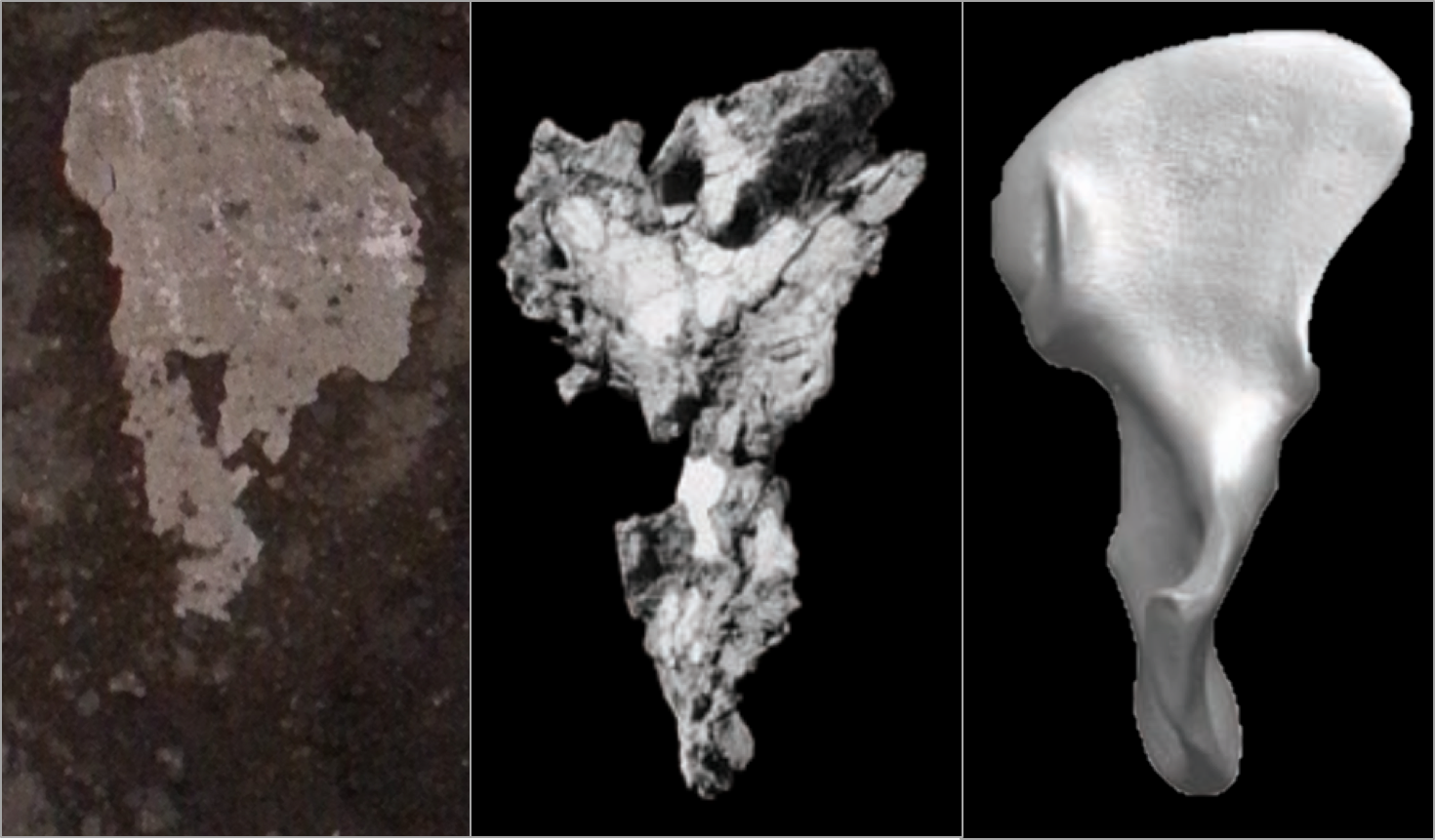
The ground section looks superficially like an unfused epiphyseal surface, whereas the rounded outer surface is more spherical. It turns out, in the most nefarious of all holiday conspiracies since the War on Christmas, nutmeg halves are nothing more than unfused femur heads! Compare with the epiphyseal surface of this Homo naledi femur head:
Nutmeg (left) and H. naledi specimen UW 101-1098 (right).
This immature H. naledi specimen was recently published (Marchi et al., in press), and the associated 3D surface scan has been available for free download on Morphosource.org for a while now. It fits onto a proximal femur fragment, UW 101-1000, also free to download from Morphosource.
Modified Fig. 11 from Marchi et al. It’s weird that only H. naledi bones were found in the Dinaledi chamber, but even weirder is the underreported presence of nutmeg.
Like most bones in the skeleton, the femur is comprised of many separate pieces that appear and fuse together at different, fairly predictable ages. The shaft of the femur appears and turns to bone before birth, and the femur head, which forms the ball in the hip joint, usually appears within the first year of life and fuses to the femur neck in adolescence (Scheuer and Black, 2000). So we know this H. naledi individual was somewhere between 1–15ish years by human standards, probably in the latter half of this large range.
So there you have it. Osteology is everywhere – the holidays are practically a pit of bones if you keep your eyes open.
REFERENCES
Marchi D, Walker CS, Wei P, Holliday TW, Churchill SE, Berger LR, & DeSilva JM (2016). The thigh and leg of Homo naledi. Journal of Human Evolution PMID: 27855981.
Scheuer L and Black S. 2000. Developmental Juvenile Osteology. New York: Elsevier Academic Press.
I went to a cafe today to eat breakfast and get some work done. Write, write, write. It’s important to be properly nourished to ensure maximal productivity.
The Ron Swanson diet.
But I was aghast to behold the food they placed before me:
What on earth is this?
First of all, this is not a sufficient amount of bacon.
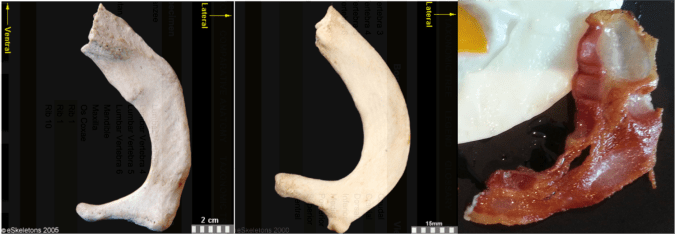
Secondably, this bacon is a spitting image of a first rib:
First ribs from the right side of the body, viewed from the top. From left to right: Human, chimpanzee, bacon. First two images from eSkeletons.org.
At the top of the ribcage, just beneath the clavicle and subclavian artery and vein, the first rib is much shorter and flatter than the rest of the ribs. As Jess Beck at Bone Broke points out, “The first and second rib give something of an awkward ‘slow song at a middle-school dance’ kind of a hug, while the lower ribs provide a more comfortable and self-assured embrace.” I mean, just lookit how sheepishly the bacon dances with the eggs in the first picture, it has ‘middle-school dance’ written all over it.
But the bacon is not totally identical to the human and chimpanzee counterparts. It’s missing their anteromedially sweeping arc, and the distal portion reaching out to the egg is fairly straight. This suggests we’re probably missing much of the original distal end. Posteriorly or dorsally (toward the bottom in the pic), it also appears to be missing much of the lateral portion including the vertebral facet. In this regard, this bacon rib looks a lot like the first rib of Homo naledi:
Full stack of ribs. Left to right: Human, bacon, Homo naledi, Dmanisi Homo erectus, Australopithecus sediba (x2), Australopithecus afarensis specimen “Lucy,” Ardipithecus ramidus, and chimpanzee. Images not to scale except Lucy and Ardi. Image credits given below.
It is hard to make good homologous comparisons among these fossils and bacon, since so many are so incomplete. But it looks like the hominins are relatively longer (front to back, or dorsoventrally) compared to the chimpanzee. That is, oriented along the rib “neck,” the ventral/distal end projects far more medially beyond the proximal vertebral facet in the chimp, while in the hominins the two ends are more flush. Ardi is really incomplete and so very hard to orient, but it may be more like the chimp (I think it needs to be rotated to the right more, to make the lateral edge more vertical like all the other specimens).
It will be interesting to see what my colleagues working on the Homo naledi thorax have to say about rib shapes and their functional importance, hopefully not too long from now.
Anyway, I really wish I had more bacon.
Fossil rib sources Dmanisi Homo erectus: Lordkipanidze D, Jashashvili T, Vekua A, Ponce de León MS, Zollikofer CP, Rightmire GP, Pontzer H, Ferring R, Oms O, Tappen M, Bukhsianidze M, Agusti J, Kahlke R, Kiladze G, Martinez-Navarro B, Mouskhelishvili A, Nioradze M, & Rook L (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449 (7160), 305-10 PMID: 17882214
Australopithecus sediba: Schmid P, Churchill SE, Nalla S, Weissen E, Carlson KJ, de Ruiter DJ, & Berger LR (2013). Mosaic morphology in the thorax of Australopithecus sediba. Science, 340 (6129) PMID: 23580537
Australopithecus afarensis and Ardipithecus ramidus: White TD, Asfaw B, Beyene Y, Haile-Selassie Y, Lovejoy CO, Suwa G, & WoldeGabriel G (2009). Ardipithecus ramidus and the paleobiology of early hominids. Science, 326 (5949), 75-86 PMID: 19810190
Portishead* came on the radio the other day, making iTunes display the cover of their album, Third. My inner osteologist rejoiced to see it prominently features a tooth!
Third album cover by Porthishead (2008). Image from Wikipedia
Well not a picture, but rather the name, of a tooth. In each quadrant of your mouth (most likely) are two premolars, commonly referred to as “bicuspids.” In the biz, we usually call these pals, “P3” and “P4.”
UW 101-1277 mandible, part of the Homo naledi holotype skull. Each capital letter stands for the tooth type (incisor, canine, premolar, and molar). Modified from Wits’ image gallery.
You might be wondering why we call them P3 and P4, when there are only two premolars per quadrant — what happened to P1 and P2? Homology to the rescue!
The ancestral condition for (eutherian) mammals was to have four premolars (and a 3rd incisor) in each side of the jaw. This is a “dental formula” of 3-1-4-3, indicating the numbers of each tooth type from front to back. Over time, different groups of animals have lost some of these teeth. Baleen whales have lost all of them.
P1 and an incisor were lost early in the evolution of Primates. Most Strepsirrhines and Platyrrhine monkeys retain this ancestral “2-1-3-3″ dental formula :
Ring tailed lemur (left) and woolly monkey (right) maxillae, showing the primitive primate dental formula including a P2. For scale, gridlines are 10 mm (left) and 20 mm (right). Images from the sadly defunct CT database from the Kyoto University Primate Research Institute.
The last common ancestor of catarrhines (living humans, apes and Catarrhine monkeys) lost the P2, and so we have only two premolars left in each side of the jaw. These are homologous with the third and fourth premolars of the earliest mammals. And that’s why we call them P3-4.
*The song was “The Rip.” It’s a very good song with an insanely creepy and trippy video:
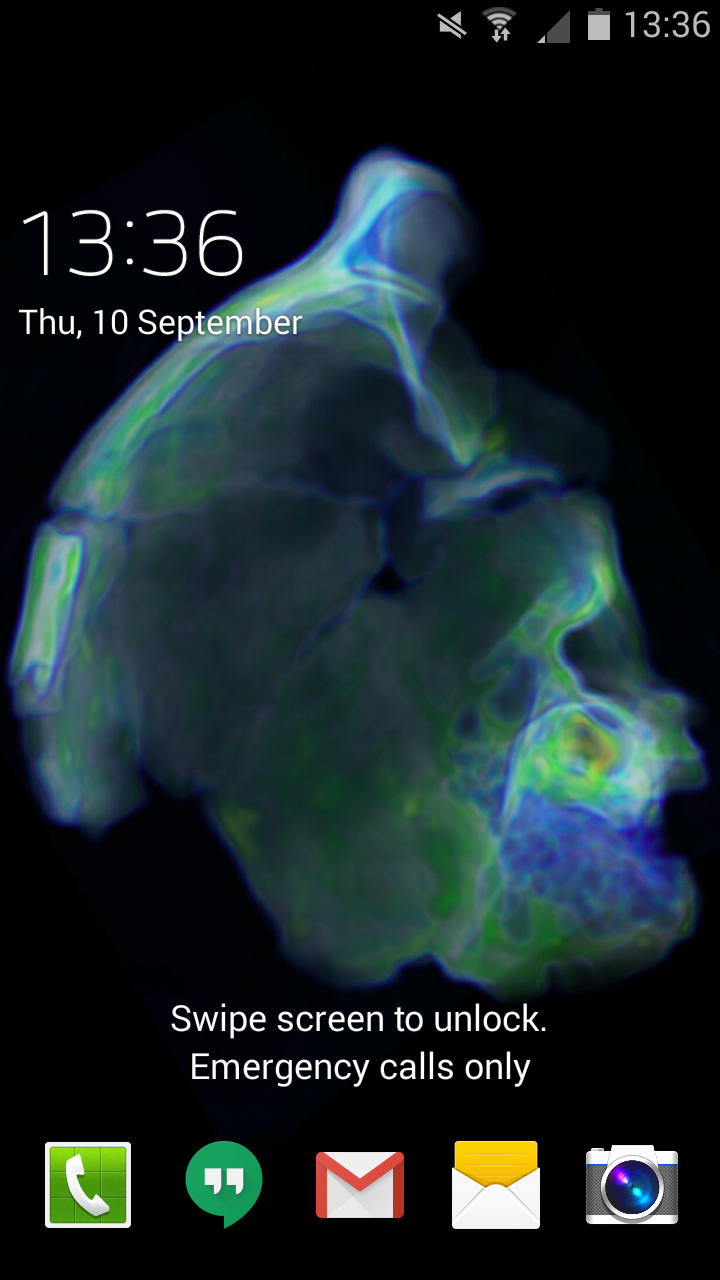
It is a great relief that Homo naledi, a most curious critter, was announced to the world on Thursday. I’ve been working on these fossils since May 2014, and it was really hard to keep my trap shut about it for over a year.
Homo naledi on my mind, and the lock screen on my phone, all year. CT rendering of cranium DH3, top is to the left and front is to the top.
I was in London for the ESHE conference last week when **it hit the fan, and so I got to attend a small press conference from the paper’s publisher, eLife, for the announcement.
eLife press conference last Thursday. From left to right: friends and colleagues Will Harcourt-Smith, Matthew Skinner, Noel Cameron, Alia Gurtov and Tracy Kivell.
I had just flown in from Kazakhstan, and was presenting some recent work on the evolution of brain growth (I’ll write a post about it soon, promise), so it was a bit hard to appreciate the gravity of the announcement. Although the awesome spread in National Geographic did help it sink in a bit.
Really blurry photo of Markus Bastir holding up the heaviest copy of National Geographic ever.
I’m wending my way back to Kazakhstan now, but in the coming weeks I will try to post more about these fossils, the project, and specifically what I’m working on.
Until then, I’d like to point out how much information is freely and easily available to the entire world about these fossils. The paper, full-length and filled with excellent images of many of the specimens and reconstructions, is available for free online here. In addition, you can download 3D surface scans of over 80 of the original fossils on MorphoSource, also totally free. Everything about this scientific discovery and its dissemination is unprecedented – the sheer number of fossils and the ease of access with which literally everyone (well, with an internet connection) can access this information has never occurred before. This is the way paleoanthropology should be. Hats off to Lee Berger and the other senior scientists on the project for making such a monumental resource available to all.
Berger LR, Hawks J, de Ruiter DJ, Churchill SE, Schmid P, Delezene LK, Kivell TL, Garvin HM, Williams SA, DeSilva JM, Skinner MM, Musiba CM, Cameron N, Holliday TW, Harcourt-Smith W, Ackermann RR, Bastir M, Bogin B, Bolter D, Brophy J, Cofran ZD, Congdon KA, Deane AS, Dembo M, Drapeau M, Elliott MC, Feuerriegel EM, Garcia-Martinez D, Green DJ, Gurtov A, Irish JD, Kruger A, Laird MF, Marchi D, Meyer MR, Nalla S, Negash EW, Orr CM, Radovcic D, Schroeder L, Scott JE, Throckmorton Z, Tocheri MW, VanSickle C, Walker CS, Wei P, & Zipfel B (2015). Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife, 4 PMID: 26354291