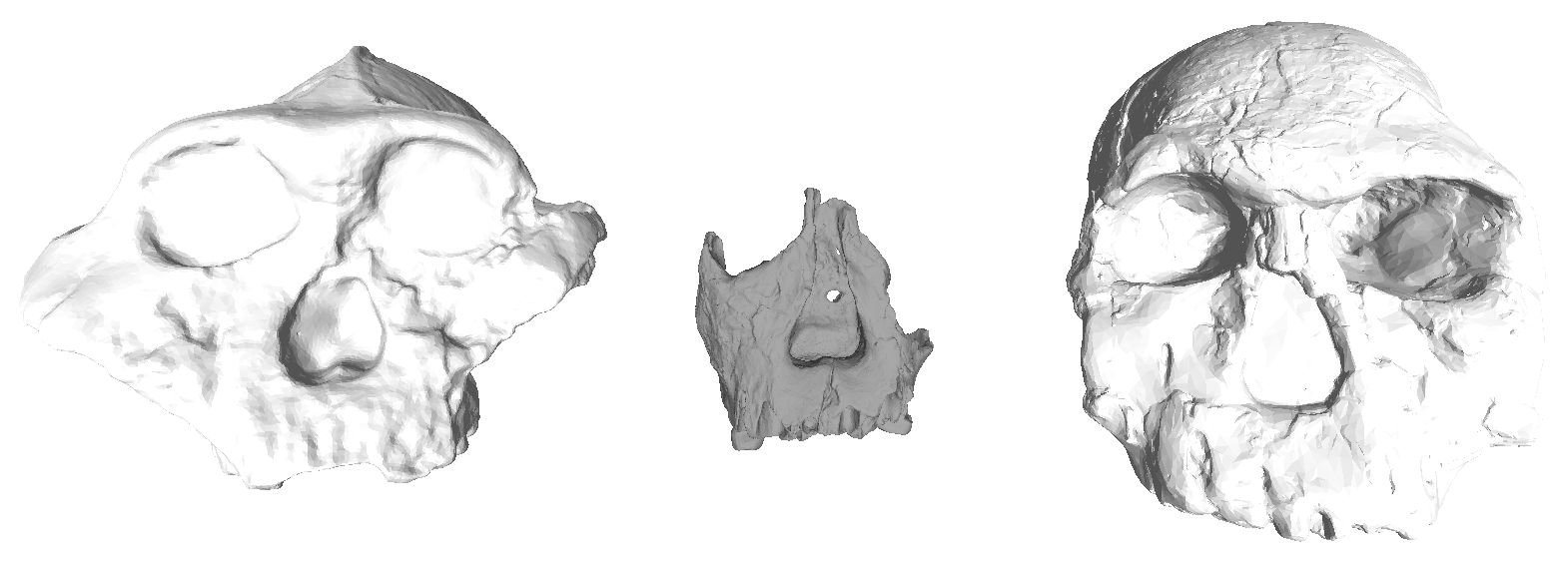
Gibbons are sometimes referred to as “lesser apes” since they’re the smaller-bodied cousins of “great apes” like us humans, chimpanzees, gorillas, and orangutans. But what they lack in body mass they make up for in taxonomic diversity, with roughly 20 species distributed across four genus groups (Kim et al., 2011). And while male great apes (except humans) have large canine teeth, both sexes in gibbons have large maxillary canines — flashy weaponry for defending territory.
Pointy canine teeth peeking out from the upper and lower jaws of an adult female gibbon cared for at the International Primate Protection League (source)
My research has generally focused on brains and growth throughout human evolution, but I started looking at gibbons a few years ago when the COVID-19 pandemic put research travel on hold. Inspired by Julia Zichello’s 2018 article about gibbon models for understanding hominin evolution and appreciating that “overlooked small apes need more attention,” I had the opportunity to CT scan a unique skeletal collection of white-handed gibbons (Hylobates lar), which was sadly harvested from the forests of Thailand back in the late 1930s. Previous research on skull growth in gibbons has mostly used small samples compiled from different species (and sometimes even different genera). In contrast, this CT dataset includes many individuals at each stage of maturation from late infancy through adulthood, effectively representing a single population at a point in time. So with this larger cross-sectional sample of a single species, we can better understand how gibbon brains and faces grow. And because permanent teeth form in a long, continuous sequence throughout the growth period, an individual’s state of dental development can serve as a marker of where they are along the maturation process.
In a paper hot off the press, Julia Boughner and I analyzed dental development in this unique sample (article here). One of the coolest things we found was that gibbons’ large upper canine teeth are among the first to begin but last to finish tooth formation. In fact, the large canines growing inside relatively small faces may inhibit growth of one of the neighboring incisor teeth until the face has grown to create enough space for it. And while most teeth developing within the jaw begin emerging into the mouth once there’s enough room for them, gibbons’ gargantuan upper canines are forced out of hiding as they outgrow their bony crypts (check out the right-most jaw in the second row below).
Cross-sectional representation of tooth formation in white-handed gibbons, starting with the youngest in the top left and ending with the oldest in the bottom right. The first permanent tooth to form and emerge, M1, is highlighted along with the canine “C.”
In addition to characterizing ‘normal’ dental development, we also observed several developmental anomalies and pathologies in the sample. Our observations corroborate previous research showing that tooth formation generally proceeds ‘as scheduled’ despite various other disturbances to development.
It remains to be seen whether early development of the canine at the cost of delayed incisor formation is a pattern unique among all the apes, since most other studies of ape tooth formation have examined the lower jaw while our study focused on the upper jaws. But the canine-incisor tradeoff that we identified sets the stage for subsequent study of skull growth in this sample, as it highlights the many factors and functions that must be coordinated during growth.
While we have several projects planned with this unique dataset, we have also published the tooth formation data that we analyzed, and the original micro-CT scans themselves will be published to the online repository Morphosource.org soon, once a few more projects are finished.
I’m recently returned from this year’s AAPA Conference, hosted by Tulane University in New Orleans. What a trip!
Usually my presentations involve fossils and/or growth, but this year I wanted to try a different way of looking at the evolution & development – integration & modularity. In short, biological structures that share a common developmental background and/or function may comprise ‘modules’ that are highly ‘integrated’ with one another, but relatively less integrated with other structures or modules.
I hypothesized that canine reduction in hominins is a result of a shift in modularity of the dentition, such that the canine became more highly integrated with the incisors than with the premolars. I’d thought of this 5 years ago when creating the first rendition of my human evo-devo course (offering again next fall!), but never got to look into it. Interestingly, the results generally supported my predictions, except for one pesky sample…
As my primatologist friends will tell you, male chimps are the worst.
Here’s a pdf version of the poster. It was fun to dabble with a new methodology, to see my far-flung friends, and to visit a fun historic place for the AAPA conference. Definitely looking forward to next year in Austin!
Today is not like the good ol’ days. In many ways things have changed for the better. For instance, in the good ol’ days, many paleontologists would find fossils but let nary a soul examine them; today, you can download high quality 3D models of many important fossils from both East and South Africa, completely for free!
Robert Broom’s (1938) account of the discovery of the first Paranthropus (or Australopithecus) robustus is also a reminder of the strangeness of the bygone days of yore:
Wait for it …
In June of this year a most important discovery was made. A schoolboy, Gert Terblanche, found in an outcrop of bone breccia near the top of a hill, a couple of miles from the Sterkfontein caves, much of the skull and lower jaw of a new type of anthropoid. Not realizing the value of the find, he damaged the specimen considerably in hammering it out of the rock. The palate with one molar tooth he gave to Mr. Barlow at Sterkfontein, from whom I obtained it. Recognizing that some of the teeth had recently been broken off, and that there must be other parts of the skull where the palate was found, I had to hunt up the schoolboy. I went to his home two miles off and found that he was at the school another two miles away, and his mother told me that he had four beautiful teeth with him. I naturally went to the school, and found the boy with four of what are perhaps the most valuable teeth in the world in his trouser pocket. He told me that there were more bits of the skull on the hillside. After school he took me to the place and I gathered every scrap I could find; and when these were later examined and cleaned and joined up, I found I had not only the nearly perfect palate with most of the teeth, but also practically the whole of the left side of the lower half of the skull and the nearly complete right lower jaw.
What a wild time – Broom hunts down poor Gert, barges into the school, then makes the kid show him where he hacked the skull out of the rock. Poor, poor Gertie.
Maybe it was a different Gertie, but surely the reaction was the same.
Of course, there was a lot at stake. I mean, brazen Gert harbored not just “beautiful teeth,” but “the most valuable teeth in the world.” IN HIS TROUSERS! And of course Gert was also the soul possessor of priceless intel – the source of the fossils. So maybe Broom was justified in this zealous abduction. And O! such prose in a Nature paper! WAS IT WORTH IT, DR. BROOM?
At Sterkfontein, a bronzed Broom considers the weight of his actions.
Of course, Gert wasn’t the last kid to discover an important human fossil. The game-changing Australopithecus sediba was discovered when Matthew Berger, son of famed Lee Berger and only 9 years old at the time, saw a piece of a clavicle sticking out of a block of breccia. Both Gert and Matthew show that you don’t have to be a doctor to make amazing discoveries. What future fossil discoveries will be made by kids, and making my adult accomplishments pale in comparison?!
After these drawings, my students are now fully trained and ready to tackle the odontological world.
. . . I’ve got dentition on the brain. WHICH IS NOT THEIR ANATOMICAL POSITION.
So last weekend some friends and I hit a local pub, a life jacket for my dental inundation. Surely, a pint and a snack will expunge enamel, dissolve dentine, exhume zuby from my brain! We ordered some beer and baursaki, delicious fried bread made out here in Kazakhstan, the perfect snack to go with beer and chechil. Tearing into the pastry, I started to feel at peace, but then was horrified to look down and find myself hoist with my own petard:
Baursak with a bite taken out? Our a hominin canine?
Seeing the snack, I saw the very thing I’d been fleeing – a hominin canine tooth. Inadvertently, I’d almost exactly replicated Sts 50, a lower left canine crown and broken root from the South African site of Sterkfontein.
Left: Sts 50, lower left canine. Right: bitten fried bread. Images not to scale. ANTIMERES?
They’re nearly identical but from opposite sides (the fancy word for which is “antimeres”). Note the tall-shouldered, sharp apex of the crown, and the little distal tubercle, the little ‘bump’ at the far left in the left picture above. The mesial, or front, crown shoulder is notably taller than the distal tubercle. At probably around 3 million years ago, Sts 50 likely belongs to Australopithecus africanus, and retains an ape-like asymmetrical crown shape compared to the more incisor-shaped canines we humans have today.
Hominin canines and definitely no cakes. Left to right: Homo baursaki, three canines from early Pleistocene South Africa, and a modern human (from White et al. 2011). Images not to scale. Note how much less asymmetrical the modern human canine crown (far right) is compared to the fossil hominins. Teeth 1, 2, 4, and 5 are from the right side while the center, Sts 50, is from the left.
Apparently all you need to go back in time is some beer and baursaki.
Holy crap 2015 was a big year for fossils. And how fortuitous that 2016 begins on a Fossil Friday – let’s recap some of last year’s major discoveries.
Homo naledi
Some Homo naledi mandibles in order from least to most worn teeth.
The Homo naledi sample is a paleoanthropologist’s dream – a new member of the genus Homo with a unique combination of traits, countless remains belonging to at leasta dozen individuals from infant to old adult, representation of pretty much the entire skeleton, and a remarkable geological context indicative of intentional disposal of the dead (but certainly not homicide, grumble grumble grumble…). The end of 2015 saw the announcement and uproar (often quite sexist) over this amazing sample. You can expect to see more, positive things about this amazing animal in 2016.
We’ll be presenting a bunch about Homo naledi at this year’s AAPA meeting in Hotlanta. I for one will be discussing dental development at Dinaledi- here’s a teaser:
As long as we’re talking about the AAPA meetings, my colleague David Pappano and I are organizing a workshop, “Using the R Programming Language for Biological Anthropology.” Details to come!
Lemur graveyard
Homo naledi wasn’t the only miraculously copious primate sample announced in 2015. Early last year scientists also reported the discovery of an “Enormous underwater fossil graveyard,” containing fairly complete remains of probably hundreds of extinct lemurs and other animals. As with Homo naledi, such a large sample will reveal lots of critical information about the biology of these extinct species.
Australopithecus deyiremeda
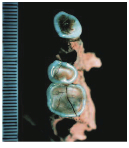
Extended Figure 1h from Haile-Selassie et al. (2015), compared with Demirjian developmental stages 6-8 . While the M1 roots look like stage 8 (complete), M2 looks like stage 7 (incomplete).
We also got a new species of australopithecus last year. Australopithecus deyiremeda had fat mandibles, a relatively short face (possibly…), and smaller teeth than in contemporaneous A. afarensis. One tantalizing thing about this discovery is that we may finally be able to put a face to the mysterious foot from Burtele, since these fossils come from nearby sites of about the same geological age. Also intriguing is the possible evidence, based on published CT images (above), that A. deyiremeda had relatively advanced canine and delayed molar development, a pattern generally attributed to Homo and not other australopithecines (if this turns out to be the case, you heard it here first!).
Lomekwian stone tool industry
3D scan and geographical location of Lomekwian tools. From africanfossils.org.
Roughly contemporaneous with A. deyiremeda, Harmand et al. (2015) report the earliest known stone tools from the 3.3 million year old site of Lomekwi 3 in Kenya. These tools are a bit cruder and much older than the erstwhile oldest tools, the Oldowan from 2.6 million years ago. These Lomekwian tools, and possible evidence for animal butchery at the 3.4 million year old Dikika site in Ethiopia (McPherron et al. 2010; Thompson et al. 2015), point to an earlier origin of lithic technology. Fossils attributed to Kenyanthropus platyops are also found at other sites at Lomekwi. With hints at hominin diversity but no direct associations between fossils and tools at this time, a lingering question is who exactly was making and using the first stone tools.
Earliest Homo
The reconstructed Ledi Geraru mandible (top left), compared with Homo naledi (top right), A. deyiremeda (bottom left), and the Uraha early Homo mandible from Malawi (bottom right). Jaws are scaled to roughly the same length from the front to back teeth; the Uraha mandible does not have an erupted third molar whereas the others do and are fully adult.
Just as Sonia Harmand and colleagues pushed back the origins of technology, Brian Villmoare et al. pushed back the origins of the genus Homo, with a 2.7 million year old mandible from Ledi Geraru in Ethiopia. This fossil is only a few hundred thousand years younger than Australopithecus afarensis fossils from the nearby site of Hadar. But the overall anatomy of the Ledi Geraru jaw is quite distinct from A. afarensis, and is much more similar to later Homo fossils (see image above). Hopefully 2016 will reveal other parts of the skeleton of whatever species this jaw belongs to, which will be critical in helping explain how and why our ancestors diverged from the australopithecines. (note that we don’t yet have a date for Homo naledi – maybe these will turn out to be older?)
Early and later Homo
Left: modified figures 2-3 from Maddux et al. (2015). Right: modified figures 7 & 13 from Ward et al. (2015). Note that in the right plot, ER 5881 femur head diameter is smaller than all other Homo except BSN 49/P27.
The earlier hominin fossil record wasn’t the only part to be shaken up. A small molar (KNM-ER 51261) and a set of associated hip bones (KNM-ER 5881) extended the lower range of size variation in Middle and Early (respectively) Pleistocene Homo. It remains to be seen whether this is due to intraspecific variation, for example sex differences, or taxonomic diversity; my money would be on the former.
Left: Penghu 1 hemi-mandible (Chang et al. 2015: Fig. 3), viewed from the outside (top) and inside (bottom). Right: Manot 1 partial cranium (Hershkovitz et al. 2015: Fig. 2), viewed from the left (top) and back (bottom).
At the later end of the fossil human spectrum, researchers also announced an archaic looking mandible dredged up from the Taiwan Straits, and a more modern-looking brain case from Israel. The Penghu 1 mandible is likely under 200,000 years old, and suggests a late survival of archaic-looking humans in East Asia. Maybe this is a fossil Denisovan, who knows? What other human fossils are waiting to be discovered from murky depths?
The Manot 1 calvaria looks very similar to Upper Paleolithic European remains, but is about 20,000 years older. At the ESHE meetings, Israel Hershkovitz actually said the brain case compares well with the Shanidar Neandertals. So wait, is it modern or archaic? As is usually the case, with more fossils come more questions.
Crazy dinosaurs
Yi qi was bringing Skeksi back, and its upper limb had a wing-like shape not seen in any other dinosaur, bird or pterosaur. There were a number of other interesting non-human fossil announcements in 2015 (see here and here), proving yet again that evolution is far more creative than your favorite monster movie makers.
What a year – new species, new tool industries, new ranges of variation! 2015 was a great year to be a paleoanthropologist, and I’ll bet 2016 has just as much excitement in store.
References (in order of appearance)
Haile-Selassie, Y., Gibert, L., Melillo, S., Ryan, T., Alene, M., Deino, A., Levin, N., Scott, G., & Saylor, B. (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity Nature, 521 (7553), 483-488 DOI: 10.1038/nature14448
Harmand, S., Lewis, J., Feibel, C., Lepre, C., Prat, S., Lenoble, A., Boës, X., Quinn, R., Brenet, M., Arroyo, A., Taylor, N., Clément, S., Daver, G., Brugal, J., Leakey, L., Mortlock, R., Wright, J., Lokorodi, S., Kirwa, C., Kent, D., & Roche, H. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521 (7552), 310-315. DOI: 10.1038/nature14464
McPherron, S., Alemseged, Z., Marean, C., Wynn, J., Reed, D., Geraads, D., Bobe, R., & Béarat, H. (2010). Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature, 466 (7308), 857-860. DOI: 10.1038/nature09248
Thompson, J., McPherron, S., Bobe, R., Reed, D., Barr, W., Wynn, J., Marean, C., Geraads, D., & Alemseged, Z. (2015). Taphonomy of fossils from the hominin-bearing deposits at Dikika, Ethiopia Journal of Human Evolution, 86, 112-135 DOI: 10.1016/j.jhevol.2015.06.013
Villmoare, B., Kimbel, W., Seyoum, C., Campisano, C., DiMaggio, E., Rowan, J., Braun, D., Arrowsmith, J., & Reed, K. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia Science, 347 (6228), 1352-1355 DOI: 10.1126/science.aaa1343
Maddux, S., Ward, C., Brown, F., Plavcan, J., & Manthi, F. (2015). A 750,000 year old hominin molar from the site of Nadung’a, West Turkana, Kenya Journal of Human Evolution, 80, 179-183 DOI: 10.1016/j.jhevol.2014.11.004
Ward, C., Feibel, C., Hammond, A., Leakey, L., Moffett, E., Plavcan, J., Skinner, M., Spoor, F., & Leakey, M. (2015). Associated ilium and femur from Koobi Fora, Kenya, and postcranial diversity in early Homo Journal of Human Evolution, 81, 48-67 DOI: 10.1016/j.jhevol.2015.01.005
Chang, C., Kaifu, Y., Takai, M., Kono, R., Grün, R., Matsu’ura, S., Kinsley, L., & Lin, L. (2015). The first archaic Homo from Taiwan Nature Communications, 6 DOI: 10.1038/ncomms7037
Hershkovitz, I., Marder, O., Ayalon, A., Bar-Matthews, M., Yasur, G., Boaretto, E., Caracuta, V., Alex, B., Frumkin, A., Goder-Goldberger, M., Gunz, P., Holloway, R., Latimer, B., Lavi, R., Matthews, A., Slon, V., Mayer, D., Berna, F., Bar-Oz, G., Yeshurun, R., May, H., Hans, M., Weber, G., & Barzilai, O. (2015). Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans Nature, 520 (7546), 216-219 DOI: 10.1038/nature14134
Portishead* came on the radio the other day, making iTunes display the cover of their album, Third. My inner osteologist rejoiced to see it prominently features a tooth!
Third album cover by Porthishead (2008). Image from Wikipedia
Well not a picture, but rather the name, of a tooth. In each quadrant of your mouth (most likely) are two premolars, commonly referred to as “bicuspids.” In the biz, we usually call these pals, “P3” and “P4.”
UW 101-1277 mandible, part of the Homo naledi holotype skull. Each capital letter stands for the tooth type (incisor, canine, premolar, and molar). Modified from Wits’ image gallery.
You might be wondering why we call them P3 and P4, when there are only two premolars per quadrant — what happened to P1 and P2? Homology to the rescue!
The ancestral condition for (eutherian) mammals was to have four premolars (and a 3rd incisor) in each side of the jaw. This is a “dental formula” of 3-1-4-3, indicating the numbers of each tooth type from front to back. Over time, different groups of animals have lost some of these teeth. Baleen whales have lost all of them.
P1 and an incisor were lost early in the evolution of Primates. Most Strepsirrhines and Platyrrhine monkeys retain this ancestral “2-1-3-3″ dental formula :
Ring tailed lemur (left) and woolly monkey (right) maxillae, showing the primitive primate dental formula including a P2. For scale, gridlines are 10 mm (left) and 20 mm (right). Images from the sadly defunct CT database from the Kyoto University Primate Research Institute.
The last common ancestor of catarrhines (living humans, apes and Catarrhine monkeys) lost the P2, and so we have only two premolars left in each side of the jaw. These are homologous with the third and fourth premolars of the earliest mammals. And that’s why we call them P3-4.
*The song was “The Rip.” It’s a very good song with an insanely creepy and trippy video:
We’re learning about the divergence between robust Australopithecus and early Homo 2.5-ish million years ago in my Human Evolution class this week. Because of this multiplicity of contemporaneous species, when scientists find new hominin fossils in Early Pleistocene sites, a preliminary question is, “What species is it?”
Scrutinizing the fossil record, asking the difficult questions. (Science credit)
To help my students learn how we know whether certain fossils belong to the same species, and to which group new fossils might belong, in this week’s lab we compared tooth sizes of Australopithecus boisei and early Homo. After seeing how tooth sizes differed between these groups, students then tested whether they could determine whether two “mystery” fossils (KNM-ER 60000 and 62000; Leakey et al. 2012) belonged either group.
Early Pleistocene hominin fossils from Kenya. Left to right: KNM-ER 406, ER 62000 and ER 1470. At the center is one f the lab’s “mystery jaws.”
Students downloaded 3D scans of hominin fossils from AfricanFossils.org, and measured buccolingual/labiolingual tooth crown diameters using MeshLab.
Early Pleistocene hominin mandibles. Left to right: KNM-ER 3230, ER 60000 (“mystery” jaw) and ER 1802.
The first purpose of this lab was to help familiarize students with skull and tooth anatomy of early Pleistocene humans. Although lectures and readings are full of images, a lab activity forces students to spend time visually examining fossils. Plus, they’re in 3D which is a whole D greater than 2D – the visual equivalent of going to eleven! The second goal of the lab was to help prepare students for their term projects, in which they must pose a research question about human evolution, generate predictions, and find and use data to test hypotheses.
If you’re interested in using or adapting this activity for your class, here are the handout and data sheet into which students enter their measurements. The data sheet specifies the fossils that can be downloaded from africanfossils.org. Some relevant fossils (i.e., KNM WT 15000 and ER 992) were not included because the 3D scans yield larger measurements than in reality.
Reference Leakey MG, Spoor F, Dean MC, Feibel CS, Antón SC, Kiarie C, & Leakey LN (2012). New fossils from Koobi Fora in northern Kenya confirm taxonomic diversity in early Homo. Nature, 488 (7410), 201-4 PMID: 22874966
It will be lots of work to prep my Human Evolution course for the Fall. This past year has seen many major fossil discoveries, and adding to the list is the newly described species Australopithecus deyiremeda (Haile-Selassie et al., 2015). The fossils come from newly announced sites in Ethiopia (here it is on a map!), dating to around 3.4 million years ago. These new fossils are contemporaneous with Australopithecus afarensis, fossils attributed to Kenyanthropus platyops, and whatever the hell the Burtele foot belongs to.
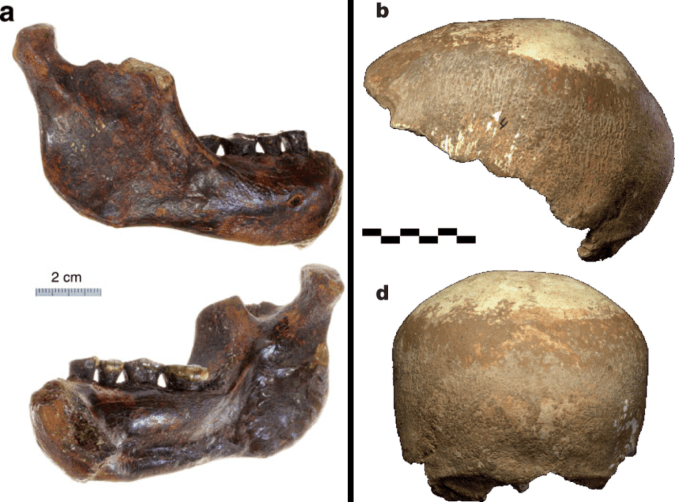
The main specimens are a fairly complete half of a maxilla (upper jaw) and two decent mandibles (lower jaw bones). These fossils do not belong to the same individual (despite all the media pictures of the upper and lower jaws together). One of the most distinctive features of these fossils is how thick, both in absolute and relative terms, the mandibles are, especially given how short they are. What sticks out to me though, is that the upper jaw looks like it might have still had some growing to do. Why on earth would I think so? (The following is based off pictures from the publications, so I could be wrong!)
Extended Figure 1a from the paper, the type specimnen BRT-VP-3/1 maxilla viewed from the left side. I’ve added the M2 label for your reading pleasure.
The holotype maxilla (BRT-VP-3/1) is described as coming from a “young adult” in the Supplementary Information. However, it looks like the second molar tooth (M2) is not quite fully erupted and in occlusion, although this could be due to the natural arc of the tooth row. There is no visible wear on the tooth in the pictures, and indeed the Supplementary Information says the tooth is unworn. This means that the tooth is only recently emerged, and may not have passed the gum line, and therefore hasn’t seen much/any use yet. Authors note in the Supplementary Information that there is no M3 (a.k.a. “wisdom tooth”) wear facet on the back of M2 , meaning the last tooth hadn’t yet emerged yet either. So, this all points to a non-adult age by tooth eruption standards.
Extended Figure 1d from the paper. Same fossil, but from the bottom; pretend you’re a dentist peering into its mouth. Back is to the bottom.
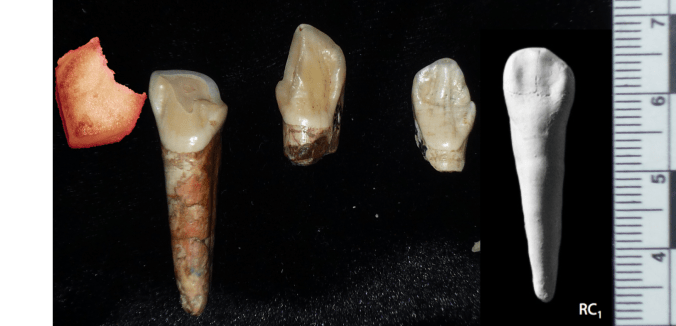
In addition, the M2 roots don’t look fully formed. This is especially apparent in Extended Figure 1h, a CT section through the teeth:
Left side: Extended Figure 1h from the paper. From left to right, the teeth are P3, P4, M1, and M2. For comparison, to the right are Demirjian tooth development stages, modified from Table 2 of Kuykendall, 1996. Also compare the M2 roots with completed roots of the M1.
In many human populations, this stage of M2 development is reached (on average) between 11-13 years (Liversidge et al., 2006). In the wild Taï Forest chimpanzee sample, two individuals with M2 root completely formed (Stage H) are 10 and 11 years old (Smith et al., 2010). These apes would not be fully mature and their facial dimensions would likely have increased had they reached adulthood (Zihlman et al., 2007).
So what this suggests to me is that this maxilla may not accurately represent adult anatomy in this newly described species. In humans, the face continues to grow downwards from adolescence into adulthood, and in apes the face continues growing both forward and downward. In the differential diagnosis of A. deyiremeda, Haile-Selassie and team state, in layman’s terms, that the cheeks are positioned more toward the front than in A. afarensis, and that the front of the face doesn’t stick out as much as in A. garhi. If this specimen was not fully grown, it is likely that the true adult anatomy would have had a face that sticks out more and has less forward-positioned cheeks than in this specimen.
But, this is all speculative, and I’d like to reiterate that these observations of dental development are based only on the published pictures. Just a thought!
Last year I brought up the implications of the small female pelvis from Gona, Ethiopia for body size variation in Homo erectus (see previous post). This individual was much smaller than other Middle Pleistocene Homo fossils, indicating size variation comparable to highly sexually dimorphic gorillas and unlike recent human populations. Before this pelvis, most known Homo erectus fossils were fairly large (comparable to living people), with only a few hints of much smaller individuals (e.g., KNM-ER 427000, KNM-OL 45500). Now joining this petite party, this tiny troop, this little lot, this compact cadre, etc., is KNM-WT 51261, a 750,000 year old molar from Kenya (Maddux et al., in press).
Occlusal area for first molars in the genus Homo. The tooth image is from Fig. 2 and the plot from Fig. 3 in Maddux et al. Lookit how tiny it is!
This ‘new’ specimen substantially increases the range of size variation among early African H. erectus molars, although the expanded range isn’t remarkable compared with later Homo samples such as from Zhoukoudian cave in China or Neandertals. What is different, though, is that most of the highly variable samples show a fairly continuous range of variation, while the WT 51261 molar is a considerable outlier from the rest of the African Middle Pleistocene sample (a lot like the situation with the Gona pelvis). So this tooth re-raises an important question: were smaller specimens like Gona and WT 51261 as rare in life as they are in the fossil record, or was such great size variation common in the Middle Pleistocene? How we reconstruct what kind of animal Homo erectus was differs depending on the answer to this question.
Two reports came out last week in the journal Nature, re-dating some European human fossils to before 40 thousand years ago (kya), a few thousand years older than previous evidence for modern-looking people in the region. The media have been reporting these studies as revealing “the first Europeans,” but of course we all know that the first Europeans were the badass hominids, my favorites, from the 1.8 million year old site of Dmanisi.
KC4 maxilla (Higham et al. 2011)
From Kent’s Cavern (United Kingdom) is a partial maxilla, now dated to 44 – 41 kya* (Higham et al. 2011; but see below). The jaw fragment with highly worn teeth was found just above some Aurignacian-like (Upper Paleolithic) blades in 1927. [NB below these 2 blades were 2 other blades of a tool “complex…tentatively associated with Neandertals] The laughable amount of bone makes it rather impossible to say whether the fossil represents a Neandertal or more modern-looking human. The authors examined what little of the morphology was left and concluded that the fossil shared the most similarities with recent humans but only a few with Neandertals. A more rigorous analysis of what this mix of traits means would have been nice (i.e. why would an individual have derived traits of both ‘modern’ humans and Neandertals?). The researchers tried to extract DNA for analysis, but apparently organic remains were too poorly preserved for a good analysis. Bummer.
Cavallo B and C (Benazzi et al. 2011)
A similar older-than-we-thought story is reported by Stefano Benazzi and buddies, who reanalyzed teeth from the Italian site of Grotta del Cavallo (left). After the site was excavated in 1967, the teeth were attributed to Neandertals and the lithics classified as “Uluzzian.” I would be a dirty liar if I said knew anything about the “Uluzzian” industry (try this other site which may be more informative), but apparently it’s seen as transitional between the Neandertal-associated Mousterian and ‘more advanced’ Upper Paleolithic toolkits. So this assemblage could be used to argue that Neandertals were smart enough to upgrade to a sexier stone tool industry shortly before their anatomy (but not their genes!) disappeared. BUT! also like in Kent’s Cavern paper, Benazzi and colleagues examined what little morphology is preserved in the fossil teeth, and (re)assigned them to modern-looking humans. The authors provided nice qualitative and quantitative arguments for the human status of the teeth, though again I have to raise caution that these are only teeth and we have no idea what the rest of the skeleton would have looked like. Researchers also analyzed shells associated with the now-human teeth and dated the site to around 44 kya, making them the oldest probably-human remains in Europe.
Now, according to conventional wisdom, the Neandertals were dullards who made and used the Mousterian stone tool industry. The Mousterian was nice and all, but not nearly as wicked-pisser as the smart and sassy modern-looking humans’ Aurignacian toolkit. The thing is, though, there really hasn’t been really a lot of evidence unequivocally linking modern-looking fossils with Aurignacian artifacts. So both of the recent studies in Nature lend support to the idea that maybe modern humans were the sole makers (and users) of an advanced stone tool industry. But it’s important to remember [1] that the blades ‘associated’ with the Kent’s Cavern jaw can’t really be definitively attributed to a stone tool industry; [2] the blades were deeper in the cave than the jaw, and so may actually be appreciably older than the jaw; and [3] while the teeth from Kent’s Cavern and Cavallo do look most comparable to those belonging to ‘anatomically’ modern humans, we don’t know what the skulls or skeletons containing the teeth looked like. All that said, it’s neat to see the possible appearance of certain anatomy and technology in Europe thousands of years earlier than previously thought. It also re-raises the issue of the degree to which modern-looking humans and Neandertals overlapped in space and time, and what these interactions would have been like (I’d guess terrifying, hilarious and/or sexy).
KC4 originally (Higham et al. 2011)
A fun lesson also comes from the Kent’s Cavern study. Higham and colleagues report that the KC4 human maxilla was excavated in 1927 and analyzed by Sir Arthur Keith, a well-trained anatomist and paleoanthropologist famous in his day. Keith described the fossil as containing a canine, second premolar and first molar (right, compare with above). Higham and colleagues, however, noticed that what Keith described as a second premolar was in fact a first premolar. The teeth are so worn they mostly lack information about their form and features, so this mistake probably didn’t really mislead anatomists in any way. Still, it goes to show that even experts like Keith can make mistakes or overlook the things they know best, and this is not the first time I’ve seen this happen with fossils. So always (politely) question those giants whose shoulders you’re standing upon.
*Update (07 Nov): John Hawks points out that the KC4 maxilla itself was not used to obtain the older radiocarbon age estimate. In 1989 the maxilla was directly dated to around 30 kya, over 10 ky younger than the new estimates. Higham and friends thought this date was too young, based on methodological grounds. An attempt to redate the KC4 maxilla based on one of the teeth yielded too little collagen (organic material) to produce a date. Bummer again! The new estimate is instead based on radiocarbon dates obtained from animal bones that were excavated from above and below the human jaw. So this ‘redating’ of the Kent’s Cavern maxilla is very interesting, but should be taken with a grain of salt.
See for yourself Benazzi, S., Douka, K., Fornai, C., Bauer, C., Kullmer, O., Svoboda, J., Pap, I., Mallegni, F., Bayle, P., Coquerelle, M., Condemi, S., Ronchitelli, A., Harvati, K., & Weber, G. (2011). Early dispersal of modern humans in Europe and implications for Neanderthal behaviour Nature DOI: 10.1038/nature10617
Higham, T., Compton, T., Stringer, C., Jacobi, R., Shapiro, B., Trinkaus, E., Chandler, B., Gröning, F., Collins, C., Hillson, S., O’Higgins, P., FitzGerald, C., & Fagan, M. (2011). The earliest evidence for anatomically modern humans in northwestern Europe Nature DOI: 10.1038/nature10484