
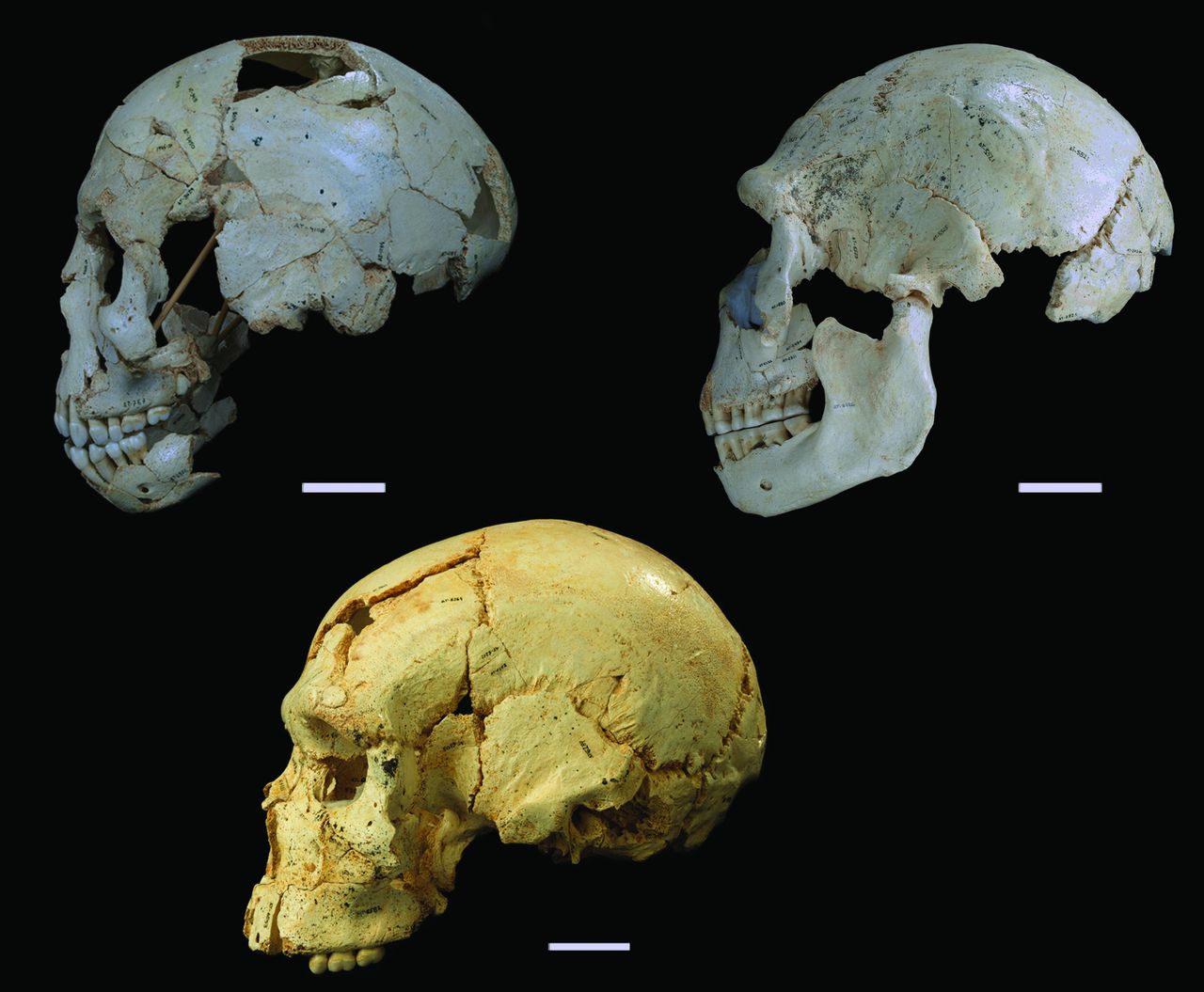
The Homo erectus infant from Mojokerto. The fossil as preserved is on the left, and on the right is the brain cast reconstructed from CT scans of the fossil (Figure 7 from Balzeau et al., 2005). The fossil and endocast are viewed from the right side so the front is on the right and back is on the left.
My paper (coauthored with Jeremy DeSilva) about brain growth in Homo erectus will be coming out soon in Journal of Human Evolution. I’ve been working on this study for a while now, so it feels good to’ve turned in the approved copy edits at long last. I’ve discussed this work a bit while it was in progress (here, here, and here), and the final version is a little different from what I posted back then, but I won’t rehash everything here. The take home message is that by around 1 million years ago, Homo erectus from Java probably had brain growth rates during early infancy in the modern human range. Really rapid early brain size growth is a unique feature of humans, and our analysis shows this trait, and many other correlates of it, were likely present early in our evolutionary history.
Our results are based on a custom resampling test, the codes for which I’ve posted here on my R Codes page. Now you can do this kind of analysis yourself!
Until the paper actually comes out, here’s the abstract:
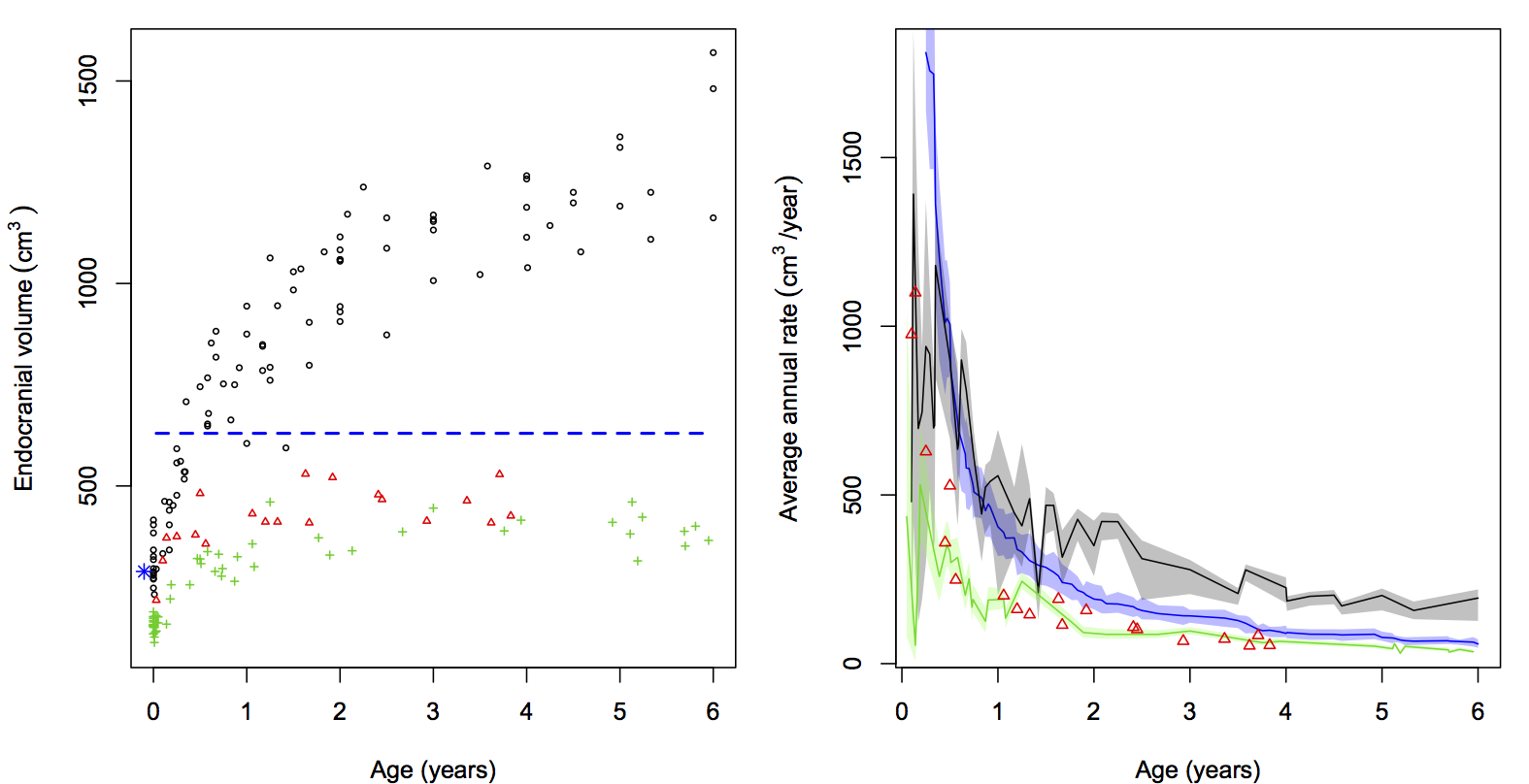
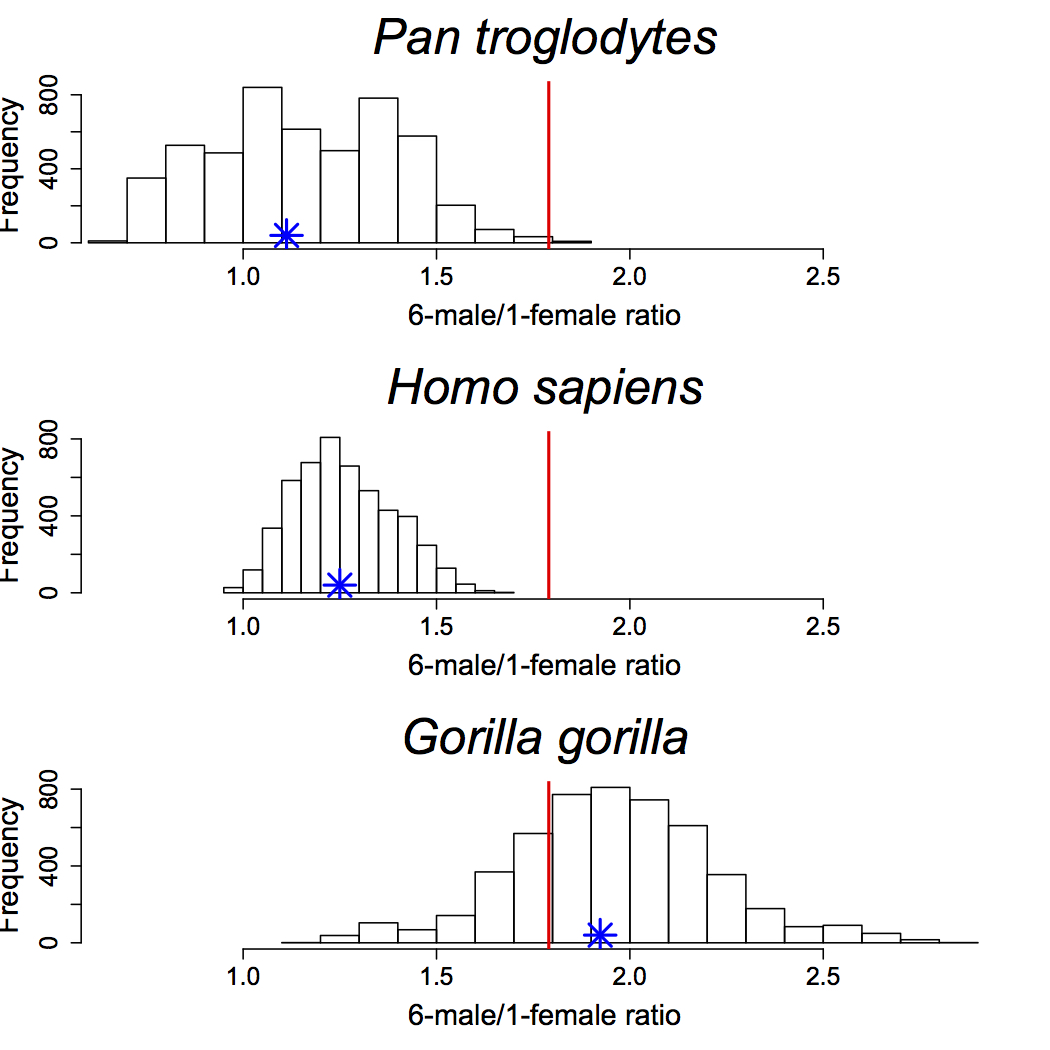
The Mojokerto calvaria has been central to assessment of brain growth in Homo erectus, but different analytical approaches and uncertainty in the specimen’s age at death have hindered consensus on the nature of H. erectus brain growth. We simulate average annual rates (AR) of absolute endocranial volume (ECV) growth and proportional size change (PSC) in H. erectus, utilizing estimates of H. erectus neonatal ECV and a range of ages for Mojokerto. These values are compared with resampled ARs and PSCs from ontogenetic series of humans, chimpanzees, and gorillas from birth to six years. Results are consistent with other studies of ECV growth in extant taxa. There is extensive overlap in PSC between all living species through the first postnatal year, with continued but lesser overlap between humans and chimpanzees to age six. Human ARs are elevated above those of apes, although there is modest overlap up to 0.50 years. Ape ARs overlap throughout the sequence, with gorillas slightly elevated over chimpanzees up to 0.50 years. Simulated H. erectus PSCs can be found in all living species by 0.50 years, and the median falls below the human and chimpanzee ranges after 2.5 years. Homo erectus ARs are elevated above those of all extant taxa prior to 0.50 years, and after two years they fall out of the human range but are still above ape ranges. A review of evidence for the age at death of Mojokerto supports an estimate of around one year, indicating absolute brain growth rates in the lower half of the human range. These results point to secondary altriciality in H. erectus, implying that key human adaptations for increasing the energy budget of females may have been established by at least 1 Ma.





 Ancient DNA was boss:
Ancient DNA was boss: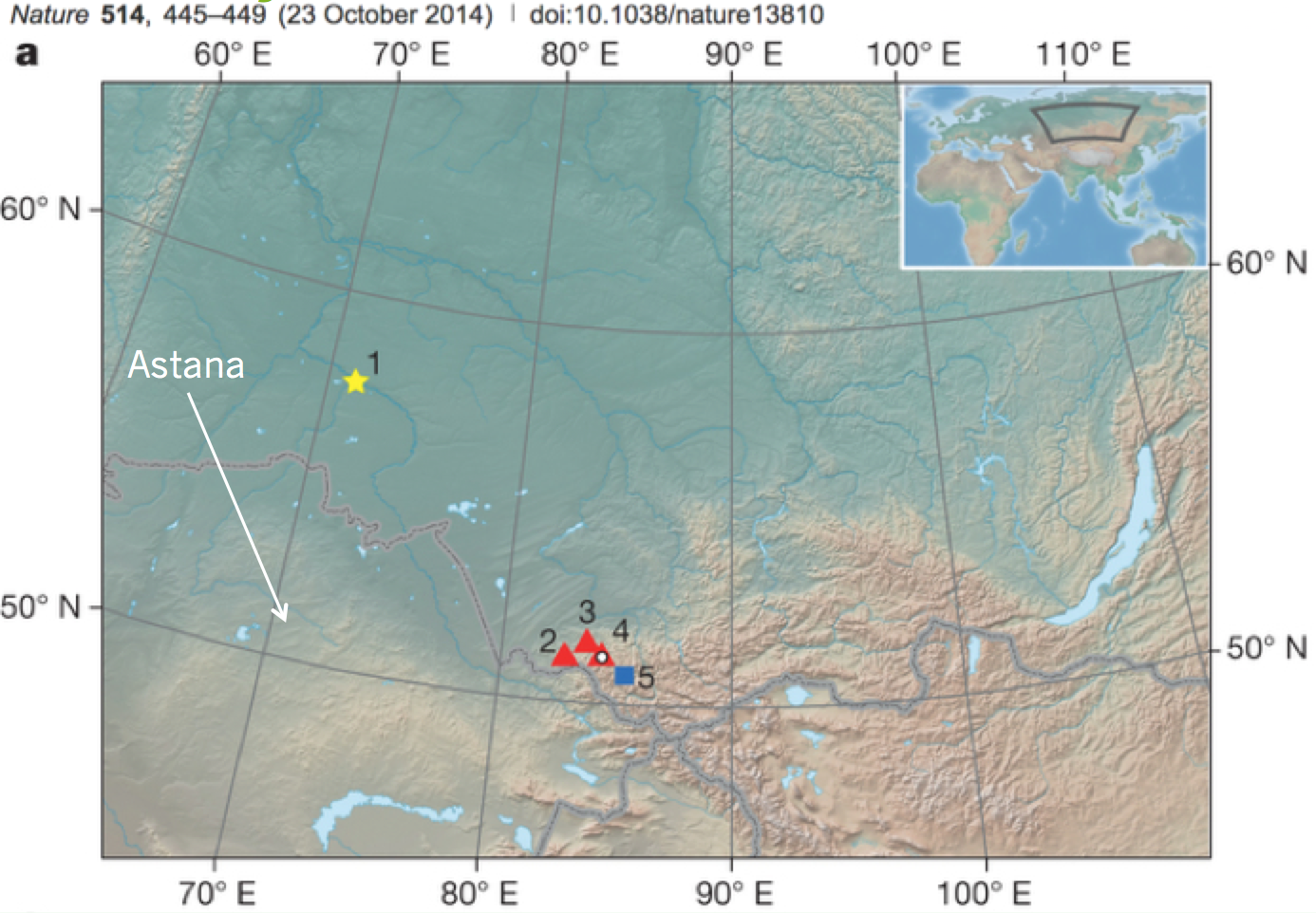











 Last week I was visiting my family in
Last week I was visiting my family in 




{kind=link}
{kind=link}