Endocasts are the faint whispers of ancient minds. These fossilized phantoms are just about all we have to tell us about the evolutionary history of brains. We are (we are, we are) rather helpless—how can we turn a whisper to a scream?
Antoine Balzeau and colleagues have recently published an incredible resource for studying brain endocasts, which they aptly call the ‘Rosetta Stone for paleoneurology.’ This is the latest paper from their project PaleoBRAIN, which draws on advanced techniques for studying endocasts and reconstructing the brains of extinct hominins. A few years ago the group published an article led by Nicole Labra asking “What do brain endocasts tell us?”, where they demonstrated the extent to which experts in brain anatomy can nevertheless misidentify actual brain impressions on endocasts. This is a big deal since the identification of brain features, namely sulci separating specific parts of the cortex, is essential for understanding how the human brain has evolved over the past several millions of years. When looking at an endocast, are we really seeing the brain structures that we think we’re seeing?
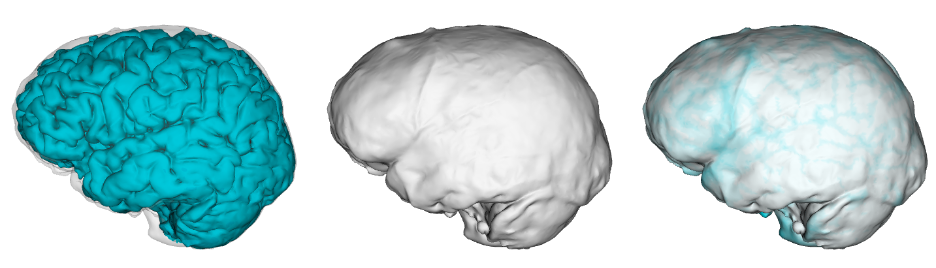
Balzeau and colleagues make another major contribution to address this problem for paleoneurology. The researchers used advanced MRI brain scanning methods to directly compare the brains and bony endocasts of 75 living humans. They used software that automatically identifies brain sulci from MRI scans and then examined the extent to which each individual’s sulci (left image, above) were expressed on their endocast (center image, above). The study expands greatly on similar work by Jean Dumoncel and colleagues using a slightly different approach. As in the previous research, Balzeau and co found that the endocast can serve as a decent proxy for the underlying brain anatomy, but with some pretty big limitations.
One of the major differences the authors identified between brain and endocast is that whereas brain sulci are often like long valleys (for instance, the long, straight lateral sulcus separating the temporal and frontal/parietal lobes), the corresponding sulci on endocasts are usually much shorter: that is, less of the sulcus makes an impression. Worse, sulci are often broken up into separate segments on the endocast. This is important because if a sulcus isn’t fully preserved we may not know its true course or the spatial relationship between certain brain structures. Plus, if a sulcus is broken up on an endocast, we risk misidentifying the different segments as other, incorrect sulci.
Perhaps the most shocking and sobering observation is that endocasts may bear imprints that are completely unrelated to any actual brain sulci, which they term “MNAS” (marks not associated with sulci). What causes these impish impressions is unclear at this point, but it raises the harrowing possibility that we might identify and interpret fossilized impressions that didn’t actually exist in the brains of ancient animals. Fortunately, Balzeau and team found that MNASes tend to be located closer to the top of an endocast where the brain is not impressing as strongly, whereas true cerebral impressions are strongest in the lower regions of the endocast.
Along these lines, one cool result of the study is that the orbitofrontal sulci, from the part of the brain sitting directly above the eye sockets, were “the most visible impressions” and were observed in all 75 of the endocasts they studied. The orbitofrontal cortex is involved in regulating emotions and impulse control (reviewed in Rudebeck & Rich, 2018), so this part of the brain may have been very important for the evolution of human social behavior. The findings of Balzeau and colleagues suggests we may be able to study this region reliably in the human fossil record. A fossil called MLD 6, for example, is best known for being a beautiful Australopithecus face (well, the right part of it). Yet the fossil is also another overlooked endocast from Makapansgat, South Africa. Specifically, MLD 6 shows pronounced impressions of several of the orbitofrontal sulci, though it is admittedly only well preserved toward the middle.

The other major contribution of this paper by Balzeau and colleagues is that all of the data are publicly available (here), meaning that other researchers can validate and expand on this research. This is huge. Historically, most paleoneurologists would have to assess a fossil endocast by consulting an atlas of brain anatomy, which overlooks normal variability. If one were lucky, they could use publications documenting brains of more than one individual, such as the annotated chimpanzee brain images published by Dean Falk and colleagues. The normal variability in both brain morphology and endocranial preservations that Balzeau and co present in this study are great resources on their own. Making all the original data available, though, is a huge step toward putting all paleoneurologists on the same page.
References
Balzeau, A., Bardinet, É., Bardo, A., Bernat, A., Derrey, T., Didier, M., Filippo, A., Hui, J., Kubicka, A. M., Labra, N., Leprince, Y., Mangin, J., Mounier, A., Prima, S., Rivière, D., Santin, M. D., & Giolland, V. (2026). The ‘Rosetta Stone’ of palaeoneurology: A detailed study of the link between the brain and the endocast on 75 volunteers. Journal of Anatomy, joa.70101. https://doi.org/10.1111/joa.70101
Cofran, Z., Hurst, S., Beaudet, A., & Zipfel, B. (2023). An overlooked Australopithecus brain endocast from Makapansgat, South Africa. Journal of Human Evolution, 178, 103346. https://doi.org/10.1016/j.jhevol.2023.103346
Dart, R. A. (1949). The cranio‐facial fragment of Australopithecus prometheus. American Journal of Physical Anthropology, 7(2), 187–214. https://doi.org/10.1002/ajpa.1330070204
Dumoncel, J., Subsol, G., Durrleman, S., Bertrand, A., De Jager, E., Oettlé, A. C., Lockhat, Z., Suleman, F. E., & Beaudet, A. (2021). Are endocasts reliable proxies for brains? A 3D quantitative comparison of the extant human brain and endocast. Journal of Anatomy, 238(2), 480–488. https://doi.org/10.1111/joa.13318
Falk, D., Zollikofer, C. P. E., Ponce de León, M., Semendeferi, K., Alatorre Warren, J. L., & Hopkins, W. D. (2018). Identification of in vivo sulci on the external surface of eight adult chimpanzee brains: Implications for interpreting early hominin endocasts. Brain, Behavior and Evolution, 91(1), 45–58. https://doi.org/10.1159/000487248
Labra, N., Mounier, A., Leprince, Y., Rivière, D., Didier, M., Bardinet, E., Santin, M. D., Mangin, J. F., Filippo, A., Albessard‐Ball, L., Beaudet, A., Broadfield, D., Bruner, E., Carlson, K. J., Cofran, Z., Falk, D., Gilissen, E., Gómez‐Robles, A., Neubauer, S., … Balzeau, A. (2024). What do brain endocasts tell us? A comparative analysis of the accuracy of sulcal identification by experts and perspectives in palaeoanthropology. Journal of Anatomy, 244(2), 274–296. https://doi.org/10.1111/joa.13966
Rudebeck, P. H., & Rich, E. L. (2018). Orbitofrontal cortex. Current Biology, 28(18), R1083–R1088. https://doi.org/10.1016/j.cub.2018.07.018
Shilton, D., Breski, M., Dor, D., & Jablonka, E. (2020). Human social evolution: Self-domestication or self-control? Frontiers in Psychology, 11. https://doi.org/10.3389/fpsyg.2020.00134