|
| The Mojokerto calvaria. You’re looking at the left side of the skull: the face would be to the left. Check it out in 3D here. |
A few months ago I posted an abridged version of the presentation I gave at this year’s meetings of the American Association of Physical Anthropologists, about brain growth in Homo erectus. This study, co-authored with Jeremy DeSilva, adopts a novel approach (see “Methods” in that earlier post) to analyze the Mojokerto fossil (right). The specimen is the only H. erectus non-adult complete enough to get a decent estimate of brain size (or rather, the overall volume of the brain case) – probably 630 to 660 cubic centimeters (Coqueugniot et al. 2004; Balzeau et al., 2004). So to study brain growth in the extinct species, we just have to connect a range of estimated brain sizes at birth (around 290 cubic centimeters, based on predictive equations by DeSilva and Lesnik, 2008) to that of Mojokerto. But, the speed of brain growth implied by this comparison depends on how old poor Mojokerto was when s/he died.
Most recently, Hélen Coqueugniot and colleagues (2004) used CT scans of the fossil to examine the fusion of its various bones, to suggest the poor kid died between six months to 1.5 years, if not even younger. Antoine Balzeau and team (2005) also studied scans of the fossil, and their analysis of its virtual endocast presented conflicting age estimates, but they argued the poor kid was probably no older than 4 years. Earlier studies had suggested the kid was up to 8 years. Now, for my previous post/conference presentation, we assumed the Coqueugniot estimate was correct – but what if we consider a full range of ages for Mojokerto, from 0.03-6.00 years?
 |
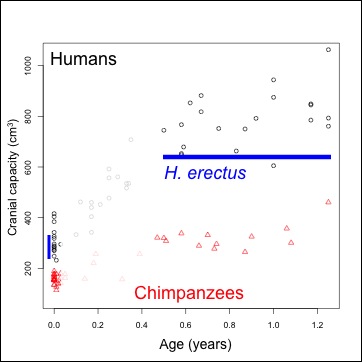
| Brain size, relative to newborns’ values, at different ages in humans (black circles) and chimpanzees (red triangles). Homo erectus median and mean are the thick solid and dashed blue lines, respectively, and the 90% and 95% confidence intervals are indicated by the thinner, dotted blue lines. Data are the same as in the previous post. |
The plot above depicts brain size relative to newborns: each circle (humans) and triangle (chimpanzees) represents the proportional size difference between a newborn (less than 1 week) and an older individual, up to 6 years. Obviously, relative brain size gets bigger in humans and chimpanzees over time. Interestingly, even though humans and chimps have very different brain sizes, the proportional brain size changes overlap a lot between species, especially at younger ages. Ah, the joys of cross-sectional samples.
But what’s especially interesting here are the blue lines on the graph, indicating estimates of proportional size change in Homo erectus, assuming Mojokerto’s skull could hold 630 cc of delicious brain matter, and that the species’ skulls at birth could hold about 290 cc, give or take several cc. The thick solid and dashed lines just above 2 on the y-axis are the mean and median of our estimates – Mojokerto’s brain averages around 2.2 times larger than predicted newborns. Such a proportion is most likely to be found in humans between 6 months to a year of age, and in chimpanzees between around 6 months and 2 years. The confidence intervals, the highest and lowest bounds of our estimates for Homo erectus proportional size change, are the thinner dashed lines on the graph. They help us constrain our estimates, and further suggest that the proportional difference found for H. erectus is most likely to be found in either chimpanzees or humans around 1 year of age – just like Coqueugniot and colleagues predicted!!!
Thus, independent evidence – brain size of Mojokerto and estimated brain size at birth in Homo erectus – corroborates a previously estimated age at death for the Mojokerto fossil, the poor little Homo erectus baby. This further supports our estimates of brain growth rates in this species, as described in the previous post.
![]() So to summarize, fairly scant fossil evidence compared with larger extant species samples using randomization statistics, argue for high, human-like infant brain growth rates in Homo erectus by around 1 million years ago. Our ancestors were badasses.
So to summarize, fairly scant fossil evidence compared with larger extant species samples using randomization statistics, argue for high, human-like infant brain growth rates in Homo erectus by around 1 million years ago. Our ancestors were badasses.
Remember, if you want the R code I wrote to do this study, just lemme know!
Those references
Balzeau A, Grimaud-Hervé D, & Jacob T (2005). Internal cranial features of the Mojokerto child fossil (East Java, Indonesia). Journal of human evolution, 48 (6), 535-53 PMID: 15927659
Coqueugniot H, Hublin JJ, Veillon F, Houët F, & Jacob T (2004). Early brain growth in Homo erectus and implications for cognitive ability. Nature, 431 (7006), 299-302 PMID: 15372030
DeSilva JM, & Lesnik JJ (2008). Brain size at birth throughout human evolution: a new method for estimating neonatal brain size in hominins. Journal of human evolution, 55 (6), 1064-74 PMID: 18789811