Last week, I introduced my Human Evolution students to the “robust” australopiths. It was a very delicate time, when we had to have a grown up, mature conversation about adult things. I reminded the students that they’re only human, but they must resist urges that seem so natural. No matter how much they want to, even if their friends are doing it, they must not act on the deep, dark desire to say that “robust” vs. “gracile” Australopithecus differ in their body build.

Don’t do it, Homo naledi. Don’t talk about body size when you mean to talk about jaw and tooth size. Illustration by Flos Vingerhoets.
Every semester, students (who don’t read and/or pay attention to lecture) think that the difference between these two groups has to do with the species’ body sizes. This is a misconception that has reached the highest echelons of reference:

Apple and Google, at least one person here is not citing their source: F-. Also, is no one else surprised that this term is allegedly specific to anthropology?
No. In the case of australopiths, “gracile” and “robust” refer to the relative size of the jaws, teeth and chewing muscles (all contributing to the “masticatory apparatus”). Traditionally, graciles include the ≥2 million year old Australopithecus afarensis and africanus, and robusts include the later A. boisei and robustus. The discovery of an A. aethiopicus cranium (Walker et al. 1986) somewhat blurred the lines between the two groups but it is usually included with the robusts (who are often collectively called Paranthropus). John Fleagle’s classic textbook (1999) illustrates the gracile-robust dichotomy very nicely:

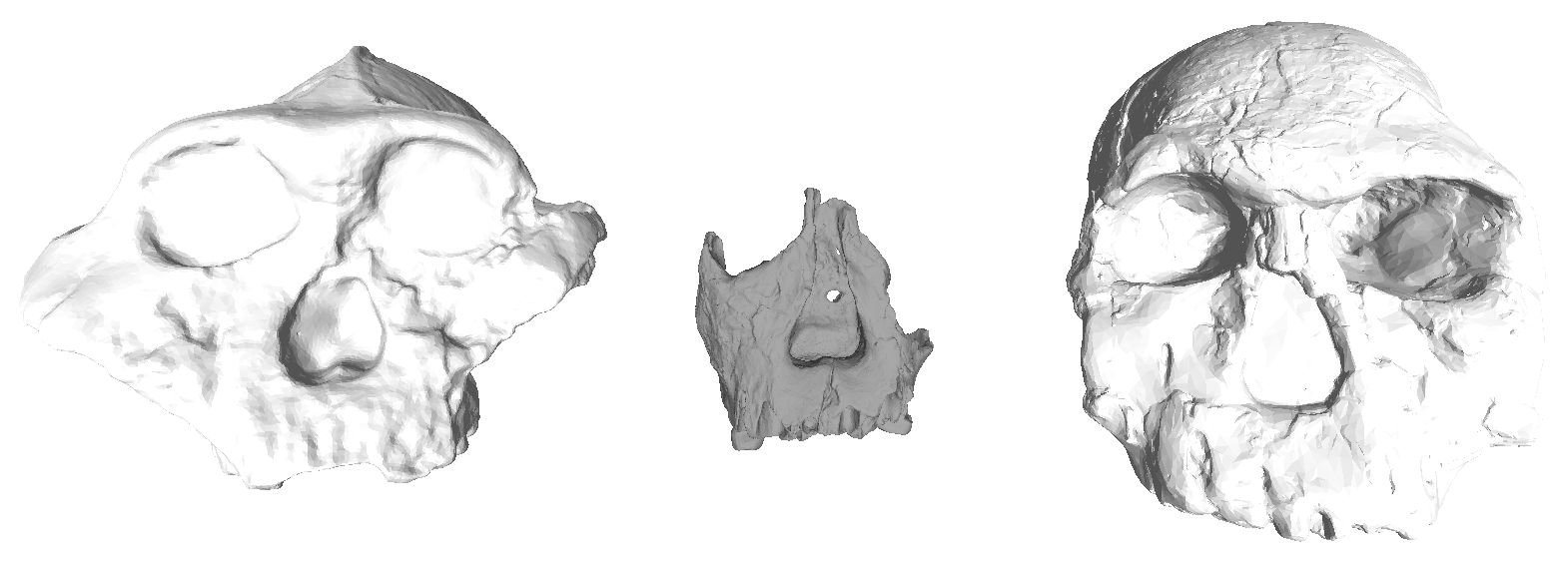
Comparison of gracile (left) and robust (right) craniodental traits. From Fleagle, 1999.
So to recap: Jaws and teeth, people! To the best of my knowledge, there’s little or no evidence that the various australopithecines differed appreciably in body size (McHenry and Coffing, 2000), stoutness, or muscularity. Although the OH 80 partial skeleton, attributed to Australopithecus boisei based on tooth size and proportions, includes a humerus with very thick cortical bone and a radius with a crazy big insertion for the biceps muscle—it was a very large and muscular A. boisei (Domínguez-Rodrigo et al., 2013). Nevertheless, gracile and robust australopithecine species differ most notably in their jaws and teeth, not bodies. Maybe this is why Liz Lemon was so confused about the term “robust”?
Today, these are somewhat antiquated terms. Back when the only early hominins known to science were the species listed above, it was easy to make a distinction. However, as the fossil record has expanded of late, the gracile-robust dichotomy becomes blurry. Australopithecus garhi (Asfaw et al., 1999) has overall tooth proportions comparable to graciles, but absolute tooth sizes and sagittal cresting like robusts. The recently described Australopithecus deyiremeda has tooth sizes and proportions like graciles but lower jaws that are very thick, like those of robust australopiths (Haile-Selassie et al., 2015).
So in light of all the confusion and blurring distinctions, maybe it’s time to scrap “gracile” vs. “robust”?
Further reading: The “robust” australopiths (Constantino, 2013). I also wrote up a short reference chapter here.

References
Asfaw B, White T, Lovejoy O, Latimer B, Simpson S, & Suwa G (1999). Australopithecus garhi: a new species of early hominid from Ethiopia. Science (New York, N.Y.), 284 (5414), 629-35 PMID: 10213683
Domínguez-Rodrigo, M., Pickering, T., Baquedano, E., Mabulla, A., Mark, D., Musiba, C., Bunn, H., Uribelarrea, D., Smith, V., Diez-Martin, F., Pérez-González, A., Sánchez, P., Santonja, M., Barboni, D., Gidna, A., Ashley, G., Yravedra, J., Heaton, J., & Arriaza, M. (2013). First Partial Skeleton of a 1.34-Million-Year-Old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania PLoS ONE, 8 (12) DOI: 10.1371/journal.pone.0080347
Haile-Selassie Y, Gibert L, Melillo SM, Ryan TM, Alene M, Deino A, Levin NE, Scott G, & Saylor BZ (2015). New species from Ethiopia further expands Middle Pliocene hominin diversity. Nature, 521 (7553), 483-8 PMID: 26017448
Walker, A., Leakey, R., Harris, J., & Brown, F. (1986). 2.5-Myr Australopithecus boisei from west of Lake Turkana, Kenya Nature, 322 (6079), 517-522 DOI: 10.1038/322517a0